














































热带农业科学(1980年创刊,月刊)第45卷第2期(总第331期)2025年2月(25日出版)目次研究论文·报告抗病特种稻新品种海农红2号选育和应用...............................唐清杰王慧艰邢福能翟李楠岑新杰严小微(1)糯米糍荔枝在莆田的引种表现及栽培技术...................................曾丽兰张游南刘小英吴碧君林革刘国强(5)野外灭绝级植物爪耳木的繁育........................................................汤欢羊青王茂媛王祝年刘咲 咲王清隆(11)不同调酸措施对酸化茶园土壤改良以及茶叶产量的影响......刘春艳赵远艳焦婷婷文勤枢胡余花熊昌云(17)海南白沙新建茶园主要害虫天敌种群与害虫消长动态分析....................................................韩芳滨于广威李超(23)硬枝黄蝉中4种环烯醚萜类化合物对辣椒疫霉菌的抑菌活性研究......................................................................................................苟亚峰田甜温思为刘世超高圣风孙世伟(33)豇豆种质资源对普通大蓟马的抗性鉴定技术规程.....................................................何云川张想芬周瀛祝增荣(37)绿农林®41号复合微生物菌肥对番茄生长影响及对青枯病防治作用...李得铭汪军翟子翔邓涛曹智醇黄俊生(43)黎药胆木根际土壤真菌群落结构分析.........................................霍天祺罗崇一杨瑞菊袁彪徐颖瑞杨卫丽(49)两种PCR快速检测转基因糜子中外源基因的插入拷贝数.........董晓静夏启玉降彦苗刘国庆程汝宏赵辉(55)激发子蛋白SbES高密度重组表达和粉剂研发杨扬陈奕鹏王茂存章淑艳刘先宝黄贵修(64)基于MAGIC群体的烟草抗黑胫病全基因组关联分析...................潘怡刘洪泰吉白璐王红黄立钰程立锐(72)基于机器学习算法的森林病虫害遥感模型对比研究......................郑绍鑫何江封成陈积标潘兴建李军集(80)金钱树种质资源倍性鉴定........................................................周成诚陆锦萍陈金花牛俊海张香秦冷青云(89)基于主成分分析法的15个秋石斛品种综合评价...........................................梁昕景李香符洁邢艳冰张孟锦(94)南宁市主城区公园绿地自生植物调查与多样性分析..........................................................................纪宝林罗应华(103)辣木固体饮料的开发及其对小鼠的减肥作用研究...........................................................吴宽赵存朝田洋鲁涛(110)虾青素益生菌生物饲料在文昌鸡中的应用...............................谯化化杨劲松谭海生窦颖鲁天宇赵松松(117)天然橡胶种植土地按园地管理的科学性及行业管理优化对策.........伍薇杨琳刘锐金何长辉刘东莫业勇(125)中国甘蔗产业区域比较优势分析.............................................................................................詹雷徐梓浩黄智刚(132)中国木薯北移的成功实践与未来展望.................................................................................................林洪鑫袁展汽(140)国家公园社区传统产业转型发展的优化路径研究——以海南热带雨林国家公园白沙茶产业为例.....谢龙缘杨奕嘉(146)综述荔枝加工技术及综合利用研究进展.....................................................................................................邓龙妹李春洪(154)[期刊基本参数] CN 46-1038/S *1980*m*A4*159*zh*P* ¥ 30.0*300*23*2025-02
CHINESE JOURNAL OF TROPICAL AGRICULTURE ISSN 1009–2196 (Monthly) Feb. 2025, Vol.45, No.2CONTENTS Research Papers and Reports Breeding and Application of the New Specialty Rice Variety Hainonghong 2 .......................................TANG Qingjie WANG Huijian XING Funeng ZHAI Linan CEN Xinjie YAN Xiaowei(1)Introduction and Performance of Nuomici Litchi (Litchi chinensis) in Putian and Its Cultivation Techniques ..................................................ZENG Lilan ZHANG Younan LIU Xiaoying WU Bijun LIN Ge LIU Guoqiang(5)Research on the Propagation of Lepisanthes unilocularis, An Extinct Plant in the Wild .............................TANG Huan YANG Qing WANG Maoyuan WANG Zhunian LIU Xiaodi WANG Qinglong(11)Effects of Different Acid-Regulating Measures on Soil Improvement and Tea Yield in Acidified Tea Plantations LIU Chunyan ZHAO Yuanyan JIAO Tingting WEN Qinshu HU Yuhua XIONG Changyun(17)Analysis of the Population Dynamics of Major Pests and Their Natural Enemies in Newly Established Tea Plantations in Baisha, Hainan ..............................................................................................................................HAN Fangbin YU Guangwei LI Chao(23)Study on the Antifungal Activity of Four Iridoids from Allemanda neriifolia Against Phytophthora capsici.............................................GOU Yafeng TIAN Tian WEN Siwei LIU Shichao GAO Shengfeng SUN Shiwei(33)Technical Procedure for the Assessment of Cowpea Germplasm Resistance to Thrips (Megalurothrips usitatus) ...................................................................................HE Yunchuan ZHANG Xiangfen ZHOU Ying ZHU Zengrong(37)Lvnong lin® 41 Compound Microbial Fertilizer on Tomato Growth and Control of Ralstonia solanacearum........................................LI Deming WANG Jun ZHAI Zixiang DENG Tao CAO Zhichun HUANG Junsheng(43)Analysis of the Rhizosphere Soil Fungal Community Structure of Liyao Nauclea officinalisHUO Tianqi LUO Chongyi YANG Ruiju YUAN Biao XU Yingrui YANG Weili(49)Rapid Detection of the Insertion Copy Number of Exogenous Genes in Transgenic Broomcorn Millet via Two PCR Methods ................................DONG Xiaojing XIA Qiyu XIANG Yanmiao LIU Guoqing CHENG Ruhong ZHAO Hui(55)High-cell-density Recombinant Expression and Powder Development of the Elicitor Protein SbES ..........................YANG Yang CHEN Yipeng WANG Maocun ZHANG Shuyan LIU Xianbao HUANG Guixiu(64)Genome-Wide Association Study for Black Shank Disease Resistance in Tobacco Based on the MAGIC Population ..........................................................PAN Yi LIU Hongtai JI Bailu WANG Hong HUANG Liyu CHENG Lirui(72)Comparative Research on Remote Sensing Models for Forest Pests and Diseases via Machine Learning Algorithms .................................................ZHENG Shaoxin HE Jiang FENG Cheng CHEN Jibiao PAN Xingjian LI Junji(80)Ploidy Identification of Zamioculcas zamiifoliaZHOU Chengcheng LU Jinping CHEN Jinhua NIU Junhai ZHANG Xiangqin LENG Qingyun(89)Comprehensive Evaluation of 15 Dendrobium hybridus Cultivars via Principal Component Analysis .......................................................................LIANG Xinjing LI Xiang FU Jie XING Yanbing ZHANG Mengjin(94)Survey and Diversity Analysis of Spontaneous Plants in the Park Green Space of the Main Urban Area of Nanning ......................................................................................................................................................JI Baolin LUO Yinghua(103)Development of Moringa oleifera Powdered Beverage and its Effects on Weight Loss .........................................................................................................WU Kuan ZHAO Cunchao TIAN Yang LU Tao(110)Application of Astaxanthin Probiotic Biological Feed in Wenchang Chickens ..................................QIAO Huahua YANG Jinsong TAN Haisheng DOU Ying LU Tianyu ZHAO Songsong(117)Scientific Analysis of the Management Model of Rubber Plantations and Optimization Strategies for Industry Management WU Wei YANG Lin LIU Ruijin HE Changhui LIU Dong MO Yeyong(125)Analysis of the Regional Comparative Advantages of China’s Sugarcane Industry .........................................................................................................................ZHAN Lei XU Zihao HUANG Zhigang(132)Successful Practice and Future Prospects for Cassava Moving Northward in China.............LIN Hongxin YUAN Zhanqi(140)Optimization Pathways for Traditional Industrial Transformation and Development in National Park Communities: A Case Study of the Baisha Tea Industry in Hainan Tropical Rainforest National Park ...............................................................................................................................................XIE Longyuan YANG Yijia(146)Review Research Progress on Processing Technology and Comprehensive Utilization of Litchi...DENG Longmei LI Chunhong(154)
2025年2月热带农业科学第45卷第2期Feb. 2025 CHINESE JOURNAL OF TROPICAL AGRICULTURE Vol.45, No.2 收稿日期 2024-02-05;修回日期 2024-03-19 基金项目 现代农业产业技术体系建设专项(No.CARS-01-92)。第一作者唐清杰(1981—),男,硕士,研究员,研究方向为热带稻类资源研究与应用,E-mail:Flyingfoxtqj@163.com。通信作者 严小微(1974—),女,本科,研究员,研究方向为水稻遗传育种,E-mail: yxwei-888@163.com。抗病特种稻新品种海农红2号选育和应用唐清杰1,2王慧艰1邢福能1,2翟李楠1岑新杰1严小微1,2(1. 海南省农业科学院三亚研究院海南三亚 572025;2. 海南省农业科学院粮食作物研究所/ 海南省农作物遗传育种重点实验室海南海口 571100)摘要海农红2号系海南省农业科学院以大丰糯作为母本、凤枣丝苗作为父本进行杂交,20142019年利用系谱选育而成的红米常规稻新品种,具有适应性广、抗病强、米质优等特点。稻瘟病和白叶枯病鉴定显示,海农红2号苗瘟抗性指数为1级高抗,白叶枯病抗性指数为3级。2020和2021年分别在海南6个市县进行区试,海农红2号抗病能力强、熟色好,具有较高的示范推广潜力。2022年通过海南省农作物品种审定委员会审定。关键词特种稻;海农红2号;选育;抗病;应用中图分类号S533 文献标识码 A DOI: 10.12008/j.issn.1009-2196.2025.02.001 Breeding and Application of the New Specialty Rice Variety Hainonghong 2 TANG Qingjie1,2 WANG Huijian1 XING Funeng1,2 ZHAI Linan1 CEN Xinjie1 YAN Xiaowei1,2(1. Sanya Institute of Hainan Academy of Agricultural Sciences,Sanya, Hainan 572025, China; 2. Cereal Crops Institute of HainanAcademy of Agricultural Sciences/Key Laboratory of Crop Genetics and Breeding of Hainan Province, Haikou, Hainan 571100, China)Abstract Hainonghong 2, a new red-rice conventional variety, was bred by the Hainan Academy of Agricultural Sciences with ‘Dafengnuo’ as the female parent and ‘Fengzaosimiao’ as the male parent from 2014 to 2019 and bred via a systematic breeding method. It features strong disease resistance, high quality, and wide adaptability. The identification shows that riceblast was grade 1, and bacterial blight was grade 3. In 2020 and 2021, Hainonghong 2 regional experiment was conducted in six counties of Hainan. The performance was strong stress resistance and disease resistance, and with good ripening color, and demonstration and promotion had high potential. It was approved by the Hainan Crop Variety Appraisal Committee in 2022. Keywords special rice; Hainonghong 2; breeding; disease resistance; application 优质、高产、抗病是水稻育种攻关的关键目标[1-2]。白叶枯病、稻瘟病是华南稻区高产、稳产的主要影响因素。选育抗病品种是比较经济、生态的措施[3-4]。目前海南水稻白叶枯研究主要集中在病菌优势生理小种[4],而抗稻瘟病、白叶枯病等育种研究较少[5-6]。人民群众越来越多关注色、香、味、营养的稻米,因此优质特种稻米可产生良好的经济效益[7]。红米是指由于色素沉积而导致种皮红色的特异水稻,前期研究主要集中于资源和选育推广,如盘州红米、福建蒲城红米、山西红香稻及陕西柳叶米[8-12]。选育和推广抗病强、产量高的优质稻,对促进海南水稻提质增效和农民增收有着重要作用[13]。海农红2号是海南省农业科学院系统选育的红米常规稻新品种,2022年通过海南省审定(审定编号琼审稻2021019)。1材料与方法1.1材料母本大丰糯,为海南省地方常规稻品种;父本凤枣丝苗(红米),为外省引进常规稻品种。1.2 方法1.2.1海农红2号选育是将大丰糯作为母本、凤枣丝苗作为父本进行杂交,利用系谱选育优良株系的方式,经8代连续选育稳定而成的常规特种红米稻。2014年晚造,用大丰糯和凤枣丝苗进行杂交,收获全部杂交种子。2015年早造,F1代种植21株,收获全部种子;当年晚造,F2代种植2 000株,选取稻米外观品质及农艺性状都比较优良的20份单株。2016年早造,F3代每个株系种植100株,选取较好性状15株;当年晚造,F4代上述15株系各种植100株,选取
2025年2月热带农业科学第45卷第2期 2 较好11株。2017年早造,F5代每个株系种植100株,选取优异9株。当年晚造,F6代每个株系种植100株,选取优良7株。2018年早造,F7代每个株系种植100株,择优选5株;当年晚造,F8代通过蒸煮品质鉴定,入选2个株系。2019年早造,参加品种比较试验,编号820,株系抗稻瘟病、白叶枯病,株型好,整齐、高产,糙米红色、光亮,按照命名顺序暂定名为海农红2号。1.2.2 农艺性状调查参照《水稻种质资源种描述规范和数据标准》[14],调查和测量有效穗数、株高、穗长、每穗总粒数、每穗实粒数、千粒重等。1.2.3 人工接种抗性鉴定稻瘟病:在水稻3~4叶期,选用当地代表性病菌喷雾接种,25~28℃为适宜接种温度;接种后遮阴网薄膜覆盖24 h,之后保湿控温;7 d后感病对照品种发病充分,调查叶瘟病级[15]。白叶枯病:水稻分蘖盛期,用剪刀蘸菌液剪去叶片顶部2 cm,接种21 d后感病对照品种发病充分,调查水稻白叶枯病病级[15]。1.2.4 米质和品质检测委托农业农村部食品质量监督检验检测中心(武汉),参照NY/T 83—1988、GB/T 21719—2008、GB/T17891—1999、GB5009.5—2010、GB/ T5009.124—2003分别进行米质、蛋白质含量、氨基酸检测[16-17]。2 结果与分析2.1 生育期海农红2号为常规稻品种,早造一般在12月至次年1月播种,生育期约130 d,晚造一般在67月播种,生育期约为120 d。20202021年,早造2年平均生育期见表1。表1 海农红2号早造生育期生育期/d 儋州琼海澄迈定安昌江海农红2号126 132 135 132 126 海农红1号(CK)126 130 133 130 124 2.2 农艺性状海农红2号分蘖力强,株型较好,谷粒细长,种皮红色。跟海农红1号相比,海农红2号平均每公顷有效穗267万(表2),平均株高105.5 cm,平均穗长23.3 cm、总粒数141.2粒、实粒数125.6粒、结实率88.9%,千粒重24.5 g。表2海农红2号农艺性状表品种穗数/(万·hm–2)株高/cm 穗长/cm总粒数/(粒·穗–1)实粒数/(粒·穗–1) 结实率/% 千粒重/g海农红2号 267 105.5 23.3 141.2 125.6 88.9 24.5 海农红1号(CK) 276 96.9 23.02 125.3 109.7 87.6 23.2 2.3 抗病性表现海农红2号苗瘟病级为1级HR,白叶枯病级为3级R;特籼占25(CK)苗瘟病级为4级,白叶枯病级为5级(表3、4)。田间种植未发现穗表3参试品种稻瘟病(苗瘟)抗性鉴定结果抗性水平品种编号HR(高抗)海黑山栏1号、海糯山栏1号、海文478s、海文486S、海农红2号MR(中抗)海香山栏 1 号,特籼占25(CK) 表4 参试品种对白叶枯病抗性鉴定结果抗性水平品种编号R(抗)海香山栏1号、海农红2号、海丰黑稻 3 号、海天黑粘1号、R225 S(感)特籼占25(CK) 颈瘟和白叶枯。2.4 米质和品质特征海农红2号米质检测结果见表5。由表5知:出糙米率77.9%,精米率67.6%,粒长6.7 mm,长宽比3.4,透明度3级,碱消值6.5级,胶稠度76 mm,直链淀粉含量13.4%,蛋白质6.92%。微量元素检测结果见表6,由表6知:铁含量9.01 mg/kg,钙含量128 mg/kg,镁含量1 108 mg/kg,钾含量2 832 mg/kg,锌含量22.0 mg/kg。17种氨基酸检测总和为5.81 g/hg,其中谷氨酸1.08 g/hg。2.5 产量和展示表现2020年早造在文昌、儋州、琼海、澄迈、定安、昌江等地初试种植。从表7可看出,平均每公顷穗数245.0万、每穗总粒数146.0、实粒数129.5、品种
唐清杰等抗病特种稻新品种海农红2号选育和应用- 3 -表5 米质检测品种出糙率/ % 精米率/ % 整精米率/% 粒长/mm粒型(长宽比)透明度(级)碱消值(级)胶稠度/mm 直链淀粉含量/% 色泽、气味蛋白质海农红2号 77.9 67.6 45.8 6.7 3.4 3 6.5 76 13.4 红米、正常6.92海农红1号(CK) 78.1 65.9 40.1 6.6 3.3 2 7 48 20.6 红米、正常8.36表6 微量物质和主要氨基酸检测结果品种铁/ (mg·kg–1) 钙/ (mg·kg–1) 镁/ (mg·kg–1) 钾/ (mg·kg–1) 锌/ (mg·kg–1) 谷氨酸/ (g·hg–1) 总氨基酸/ (g·hg–1) 海农红2号 9.01 128 1108 2832 22 1.08 5.81 海农红1号(CK) 11 143 1186 2813 24.1 1.24 6.69 表7 海农红2号产量性状2020年品种/地点穗数/ (万·hm–2) 总粒数/ (粒·穗–1) 实粒数/ (粒·穗–1) 结实率/% 千粒重/g 理论产量/ (t·hm–2) 实割产量/(t·hm–2) 海农红1号(CK) 260.5 142.5 126.5 88.8 21.5 7.08 6.54 琼海247.5 145.3 130.8 90.0 24.2 7.83 6.75 儋州244.5 148.5 130.6 87.9 24.0 7.66 7.12 文昌240.0 148.2 130.2 87.9 24.0 7.50 6.90 昌江267 148.0 128.5 86.8 23.7 8.13 7.24 澄迈247.5 140.5 128.6 91.5 24.3 7.73 6.91 定安252.0 145.5 128.5 88.3 23.6 7.64 6.90 平均245.0 146.0 129.5 88.7 23.9 7.58 6.97 2021年品种/地点穗数/ (万·hm–2) 总粒数/ (粒·穗–1) 实粒数/ (粒·穗–1) 结实率/% 千粒重/g 理论产量/ (t·hm–2) 实割产量/(t·hm–2) 海农红1号(CK) 261.5 141.7 126.8 89.5 21.7 7.20 6.57 琼海243.0 143.2 130.0 90.7 24.0 7.58 6.93 儋州249.0 145.2 128.5 88.5 23.7 7.58 6.91 文昌249.0 140.5 126.0 89.7 24.0 7.53 6.85 昌江262.5 146.5 127.8 87.2 23.0 7.72 7.11 澄迈244.5 143.8 132.6 92.2 24.5 7.94 7.19 定安247.5 146.6 130.6 89.1 23.2 7.50 6.76 平均249.3 144.3 129.3 89.6 23.7 7.64 6.90 结实率88.7%、千粒重23.9 g,每公顷实际收割产量为6.97 t,比海农红1号(CK)增产6.57%。2020年5月,相关专家在琼海大路镇对海农红2号进行现场评议:谷粒长粒,稻米外观种皮红色,田间未发现病虫危害。海农红2号机械收割1.2亩(1亩≈667 m2),平均产量450.3 kg/亩。而海农红1号(CK)机械收割1.1亩,平均产量433.4 kg/亩。2021年早造在文昌、琼海、儋州、澄迈、昌江、定安等复种展示。从表7看,平均每公顷穗数249.3万,每穗总粒数144.3、实粒数129.3、结实率89.6%,千粒重23.7 g,平均每公顷实际收割产量为6.90 t,比海农红1号(CK)增产5.63%。2021年5月相关专家在澄迈县永发镇对海农红2号进行现场复议:海农红2号机械收割1.0亩,平均产量479.1 kg/亩。而海农红1号(CK)机械收割1.0亩,平均产量454.0 kg/亩。
2025年2月热带农业科学第45卷第2期 4 3 结论大多数红米品种在我国不同地域条件下表现出很多变异类型,缺点是口感普遍较差;而且植株通常较高,遇到台风暴雨时容易倒伏,导致产量较低[18]。与普通的白米相比,红米营养丰富,是人体补充营养的佳品,更因海南红米产量低(亩产250 kg左右)市场上供不应求,使其显得十分的珍贵。海农红2号口感软粘,产量比普通红米高出近一倍。综合2年海南6个市(县)的种植表现,海农红2号抗逆性和抗病性强、不倒伏、后期熟色较好、产量较高,具有较好的示范推广潜力,建议进一步扩大示范种植。参考文献[1] 何秀英, 廖耀平, 程永盛, 等. 水稻品质研究进展与展望[J]. 广东农业科学, 2009, 36(1): 11-16. [2] 何秀英, 廖耀平, 陈钊明, 等. 优质抗病水稻新品种粤农丝苗的选育及应用[J]. 中国稻米, 2014, 20(2): 69-70. [3] 汪文娟, 周继勇, 汪聪颖, 等. 八个抗稻瘟病基因在华南籼型杂交水稻中的分布[J]. 中国水稻科学, 2017, 31(3): 299-306. [4] 夏立琼, 李明容, 谢仕猛, 等. 海南水稻白叶枯病菌优势生理小种的分离及致病力分析[J]. 分子植物育种, 2016, 14(5): 1 336-1 340. [5] 唐清杰, 韩义胜, 严小微. 多抗基因聚合的特种稻海丰黑糯2号抗病虫鉴定和分子标记检测[J]. 分子植物育种, 2018, 16(2): 466-471. [6] 唐清杰, 严小微, 唐力琼, 等. 海南地方稻新种质抗病虫鉴定与评价[J]. 中国稻米, 2019, 25(2): 50-52+58. [7] 韩龙植, 南钟浩, 全东兴, 等. 特种稻种质创新与营养特性评价[J]. 植物遗传资源学报, 2003, 4(3): 207-213. [8] 郑兴飞, 董华林, 高艳琼, 等. 我国红米资源研究进展与开发前景[J]. 农业科技通讯, 2019, (6): 4-6. [9] 黎毛毛, 余丽琴, 付军如, 等. 江西红米稻种资源主要农艺性状及营养特性分析与评价[J]. 植物遗传资源学报, 2008, 9(4): 5. [10] 杨忠义, 曹永生, 苏艳, 等. 云南作物资源特征特性及生态地理分布研究I. 云南地方稻种资源中特种稻资源[J]. 植物遗传资源学报, 2006(3): 331-337. [11] 游俊梅, 陈惠查, 金桃叶, 等. 贵州有色稻米种质资源的评价和利用[J]. 种子, 1998(4): 52-54+67. [12] 师江, 李倩, 李维峰, 等. 不同产地紫米营养成分比较及其相关性分析[J]. 热带作物学报, 2022, 43(11): 2 324-2 333. [13] 司徒志谋, 罗森辉, 蔡惠娇. 广东省优质水稻生产现状与发展策略[J]. 中国稻米, 2004, 10(2): 41-42. [14] 韩龙植, 魏兴华, 曹桂兰. 水稻种质资源描述规范和数据标准[M]. 北京: 中国农业出版社: 66-69. [15] 朱小源, 曾列先, 高汉亮, 等. 华南籼稻新品种抗病性鉴定及筛选[J]. 广东农业科学, 2010, 37(9): 12-15. [16] 唐清杰, 王惠艰, 严小微. 特种稻品种海丰黑糯2号的选育及栽培技术要点[J]. 中国稻米, 2018, 24(1): 111-112. [17] 杨国峰, 黄春燕, 王波, 等. 海南山栏稻稻米品质分析及优异资源筛选[J]. 植物遗传资源学报, 2017, 18(1): 40-45. [18] 武文豪, 何冲冲, 王传波, 等. 特种稻黑米和红米研究进展[[J/OL]]. 植物遗传资源学报, (2024-02-02)[2024-03-03] https://doi.org/10.13430/j. cnki.jpgr.20231224002. (责任编辑龙娅丽)
2025年2月热带农业科学第45卷第2期Feb. 2025 CHINESE JOURNAL OF TROPICAL AGRICULTURE Vol.45, No.2 收稿日期 2024-03-22;修回日期 2024-12-19 基金项目 福建省科技计划项目“荔枝种质资源收集、评价及应用研究”(No. 2021N0038)。第一作者曾丽兰(1988—),女,硕士,农艺师,研究方向为果树育种与栽培技术,E-mail:18050590960@163.com。通信作者 刘国强(1966—),男,本科,研究员,研究方向为果树育种与栽培技术,E-mail:liugq66@163.com。糯米糍荔枝在莆田的引种表现及栽培技术曾丽兰张游南刘小英吴碧君林革刘国强(莆田市农业科学研究所福建莆田 351106)摘要为调整优化莆田荔枝品种结构,莆田市农业科学研究所引进糯米糍荔枝在莆田试种,对其在莆田的物候期、生长情况和果实性状进行了观测分析,同时总结糯米糍荔枝的关键栽培技术要点,包括建园、施肥、树体管理、防裂果、病虫害防控和采收等。20212023年连续3年的调查结果表明,该品种在莆田的综合性状表现较好,果实平均单果重25.22 g,可食率76.64%,可溶性固形物20.30%,可滴定酸含量0.21%,优于当地主栽品种陈紫,较陈紫晚熟约13 d,较适宜在山地栽培。引进推广糯米糍荔枝可以扩大莆田荔枝种植区域,调整优化莆田荔枝品种结构。关键词糯米糍荔枝;莆田;引种;表现;栽培技术中图分类号S667.1 文献标识码 A DOI: 10.12008/j.issn.1009-2196.2025.02.002 Introduction and Performance of Nuomici Litchi (Litchi chinensis) in Putian and Its Cultivation Techniques ZENG Lilan ZHANG Younan LIU Xiaoying WU Bijun LIN Ge LIU Guoqiang (Putian Institute of Agricultural Sciences, Putian, Fujian 351106, China) Abstract To adjust and optimize the variety structure of Putian litchi, the Putian Institute of Agricultural Sciences introduced ‘Nuomici’ Litchi for trial planting in Putian, observed and analyzed phenological phases, tree growth, fruit characteristics in Putian, and summarized the key cultivation technology points of ‘Nuomici’ Litchi, including garden construction, fertilization, tree management, prevention and control of fruit cracking and control, pest control and harvesting, etc. From 2021 to 2023, the three-year observations revealed that ‘Nuomici’ Litchi had better comprehensive performance, the average fruit weight of ‘Nuomici’ was 25.22 g, the edible rate was 76.64%, the soluble solids content was 20.30%, the titratable acid content was 0.21%, which was better than the local main variety ‘Chenzi’. ‘Nuomici’ Litchi was 13 days late compared to the local ‘Chenzi’, which was more suitable for cultivation in mountainous areas. Introducing and popularizing ‘Nuomici’ litchi can expand the planting area of Putian litchi and adjust and optimize the variety structure of Putian litchi. Keywords ‘Nuomici’ litchi (Litchi chinensis); Putian; introduction; performance; cultivation techniques 荔枝Litchi chinensis Sonn.为无患子科荔枝属常绿果树,果实营养丰富,是一种食疗俱佳的南方特色水果,深受消费者的青睐[1]。糯米糍荔枝,古称水晶丸,素有荔枝之王之称,据《岭南荔枝谱》记载,该品种首见于广州萝岗街玉岩书院,在我国已有千年的栽培历史,其以色泽美、味道甜、口感佳且营养高而闻名,现已成为广东、广西、海南等地的主栽品种,种植效益明显高于一般品种,与增城挂绿、桂味并称为中国荔枝三剑客[2-4]。莆田市地处我国东南部的福建省中部沿海,荔枝是莆田四大名果之一,莆田荔枝种植历史久远,始于唐代,但目前莆田荔枝品种结构较单一、种植规模较小,荔枝种植面积仅有0.03万hm2,总产量0.12万t[5],主栽荔枝品种以陈紫为主,该品种主要分布在莆田市延寿溪、木兰溪两岸及部分乡镇沟河两畔水源丰富、阳光充足处,适宜栽培区域较狭窄,制约了莆田荔枝产业的发展。为筛选出适宜莆田山地栽培的优质荔枝品种,扩大莆田荔枝种植区域,2016年莆田市农业科学研究所引进糯米糍荔枝,接穗高接在10年生陈紫荔枝上,经多年观察其结果表现良好。现将糯米糍荔枝在莆田的引种表现及配套栽培技术要点总结如下。
2025年2月热带农业科学第45卷第2期 6 1 材料与方法1.1 引种地引种地位于福建省莆田市城厢区龙桥街道北磨社区莆田市农业科学研究所凤凰山果树基地荔枝种质资源圃,东经118°9947,北纬25°4403,属海洋性亚热带季风气候,海拔85 m,年均温在18~21℃,年日照时数1 995.9 h,无霜期300~350 d,绝对低温在0℃以上,年降雨量1 000~1 800 mm,土质疏松,土层深厚,排灌良好,正南向山坡地。土壤为红壤土,pH 6.1,有机质含量19.26 g/kg,碱解氮含量79.18 mg/kg,有效磷含量8.57 mg/kg,有效钾含量168.21 mg/kg,有效锌含量6.13 mg/kg。1.2方法1.2.1品种引进引种品种为糯米糍荔枝,于2016年10月从广东引入莆田市农业科学研究所凤凰山果树基地(福建省莆田市城厢区龙桥街道北磨社区),以10年生的陈紫荔枝为砧木,通过低位嫁接切接法进行引种试验。对照材料为同一区域的陈紫荔枝。株行距4 m×5 m,土肥水管理基本一致。1.2.2 项目调查(1)植物学特征观测在引种基地分别随机选取10株糯米糍和陈紫荔枝进行挂牌和调查,调查植物学特征[6]、病虫害、枝梢抽发、花果发育及果实成熟期等关键物候期[7]。花期隔天观察1次,其他时期每隔3~10 d观测1次。干周为嫁接口上5 cm处树干周长。(2)果实性状分析在果实成熟期从挂牌的10株树上随机选取成熟果实30个,观察果实外观特性,单果重选用英衡电子天平测定,纵径、横径、侧径、果肉厚度和果皮厚度用三丰数显游标卡尺测定;可溶性固形物含量、可滴定酸含量用Atago PAL-1数显折射仪测定,固酸比=可溶性固形物含量/可滴定酸含量。1.2.3 数据分析采用 Excel 2020和SPSS 27.0软件进行数据统计与分析。2结果与分析2.1 植物学特征糯米糍荔枝为常绿高大乔木,树势壮旺,树冠宽阔高大呈半圆形或伞形,树姿开张。6年生高接树树高3.1~4.0 m,干周31~36 cm,平均冠幅3.5 m×3.3 m,枝条细而分枝较多;一年生枝梢长度13~17 cm,叶披针形,叶长7.9~10.1 cm,叶宽3.6~4.1 cm,叶尖渐尖而歪,叶缘波浪形;小叶为椭圆形,平面互生,2~3对,小叶柄颜色为浅绿色,绿色叶面, 叶背为灰绿色,主脉为黄绿色, 侧脉不明显;树干灰褐色,树干表面光滑;花序为复总状长圆锥形, 花序中等大,雌雄同穗异花,花穗长,颜色为淡黄色,花量较多,为500~700朵,花枝细小而密。陈紫树冠半圆头形,树势强,树体高大。6年生高接树树高2.8~3.8 m,干周29~34 cm,平均冠幅3.7 m×3.2 m;一年生枝梢长度15~18 cm,叶片长披针形,叶长8.4~11.3 cm,叶宽2.7~3.1 cm,叶尖尾尖,叶面较平,叶缘微波浪形,成熟叶色浓绿;小叶对数3~4对,叶片对生;长穗状花序,花量大,但有“爱花不惜子”之称。2.2 生长结果习性糯米糍荔枝一年抽梢4~5次,春梢在3月上旬萌动, 夏梢在5月上旬萌动, 秋梢在7月底萌动,冬梢在暖冬时易抽生。糯米糍结果树一般萌发2次秋梢,7月下旬至8月上旬抽发第一次秋梢,此次抽梢有利于恢复树势和培养第二次秋梢;9月下旬至10月上旬抽发第二次秋梢以培养成第二年的结果母枝,利于次年开花结果[8]。自2016年引进糯米糍荔枝以来,该品种已在莆田推广种植面积达115亩(1亩≈667 m2),经多年观察,其在嫁接后3~5年可投产,第5年进入丰产期,2021—2023年连续3年对糯米糍和陈紫荔枝进行现场测产,糯米糍3年平均单株产量分别为23.12、26.33、32.73 kg,陈紫平均单株产量为8.9、13.53、16.73 kg,糯米糍较为丰产。2.3 物候期在福建莆田,糯米糍荔枝的花芽诱导期始于12月中旬,翌年1月下旬为花穗露白点期, 抽穗期为2月下旬,初花期4月上中旬,盛花期4月中下旬,谢花期4月下旬至5月上旬,花期约20 d,初始坐果期5月上中旬;糯米糍荔枝属于易落果品种,共有4次生理落果高峰,第一次生理落果高峰期5月中旬,第二次生理落果高峰期5月下旬,第三次生理落果高峰期6月上旬,果实着色期6月下旬,第四次生理落果高峰期在采前的7月上旬;由谢花至果实成熟80~90 d,果实成熟期7月中下旬,较主栽品种陈紫晚熟约13 d(表1),有利于延长莆田荔枝的销售供应期。
曾丽兰等糯米糍荔枝在莆田的引种表现及栽培技术- 7 -表1 糯米糍和陈紫荔枝品种在福建莆田的物候期比较品种花穗露白点期抽穗期初花期盛花期谢花期初始着果期第一次生理落果期第二次生理落果期果实着色期采前落果期果实成熟期糯米糍1月下旬2月下旬4月上中旬4月中下旬4月下旬至5月上旬5月上中旬5月中旬5月下旬至6月上旬6月下旬7月上旬7月中下旬陈紫1月中旬2月中旬4月上旬4月中旬4月下旬5月上旬5月上旬5月下旬6月中旬6月下旬7月上中旬2.4 果实主要经济性状综合糯米糍荔枝的结果数据(表2、3),与当地主栽品种陈紫荔枝相比,果实形状扁心形,果肩一边明显隆起,果顶浑圆, 果皮暗红色,龟裂片较大且平滑,乳突状隆起呈狭长型,裂片峰锐尖, 缝合线较明显;果肉同为乳白色,内膜褐色较少,质细嫩爽脆,不易流汁,风味浓甜,带蜜香;种子小,多退化或中空,焦核率显著高于陈紫,达到90%。单果重比陈紫高1.92 g,果实纵径、横径和侧径均显著高于陈紫。果皮厚度差异不显著,果肉厚度、可食率和可溶性固形物含量均显著高于陈紫,可滴定酸含量比陈紫低0.13%,固酸比比陈紫高44.9。结果说明,糯米糍荔枝果实综合性状表现较好,在福建莆田地区可进行推广。表2 糯米糍和陈紫荔枝品种在福建莆田的果实特征比较品种果实形状果皮颜色果肩形状果顶形状龟裂片形态龟裂片片峰形态缝合线果肉颜色果肉内膜褐色果肉质地流汁情况风味香味糯米糍扁心形暗红一平一隆起浑圆乳突状隆起锐尖明显乳白色少细嫩爽脆不易流汁浓甜蜜香陈紫心形或卵圆形鲜红平浑圆锥尖状突起细尖不明显乳白色中细嫩软滑易流汁浓甜微酸微香表3糯米糍和陈紫荔枝品种在福建莆田的果实性状比较品种单果重/g 果实纵径/mm 果实横径/mm 果实侧径/mm果肉厚度/mm 果皮厚度/mm糯米糍 (25.22±1.79) a (36.53±1.70) a (36.48±1.72) a (34.47±1.75) a (13.53±0.89) a (0.83±0.14) a陈紫 (23.3±1.13) b (34.22±1.12) b (31.37±1.42) b (32.37±1.42) b (10.51±0.9) b (0.84±0.11) a品种可食率/% 可溶性固形物含量/%可滴定酸含量/% 固酸比焦核率/% 糯米糍 (76.64±1.82) a (20.30±1.09) a (0.21±0.03) b 96.35 90 陈紫 (71.22±1.21) b (17.47±0.91) b (0.34±0.03) a 51.45 20 注:同列不同小写字母表示差异显著(p<0.05)。3 栽培技术要点3.1 园地选择糯米糍荔枝是“高需冷型”果树,其花芽分化需要适度的低温(冬天日平均气温11~14℃),低温有利于产生较高质量的花穗,但气温过低易产生冻害[9]。莆田大多年份气温最低月份为1~2月,平均气温为12℃左右,无严寒,温度很适宜糯米糍荔枝生长[10]。选择在地势向阳、水源充足、排灌良好、土层深厚、土壤保水保肥能力强、土质疏松肥沃、无霜冻的地块建园。因莆田地区年降雨量较大、台风较多,应建设“外埂内沟”等高环山梯田以防水土流失,即台面宽度≥3.5 m,且稍向内斜,以石块砌梯壁,在每级梯田外沿筑土埂(高度15~20 cm),内侧建排灌水沟,并在园区内建一个蓄水池便于灌溉。3.2低位嫁接方法选取树势良好且无病虫害的陈紫荔枝作为砧木,于春季进行重回缩修剪,锯断大枝时注意锯口要平整光滑且略微倾斜(防止积水)、树皮完整无扯裂,留下1~2支带叶枝条作抽水枝,留抽
2025年2月热带农业科学第45卷第2期 8 水枝有利于提高嫁接成活率。选取芽眼饱满、健壮、无病虫害、平直的1~2年生糯米糍荔枝枝条作为接穗。采用低位嫁接切接法,即选择在主干上嫁接,该方法有利于输送营养,能快速恢复树势,且促进矮化控冠,从而节省用工成本。嫁接时期宜选择在春季或早秋,避开低温、潮湿阴雨、高温暴晒等天气。嫁接时要注意快、准、严,嫁接速度要快,在切砧木部位时要注意根据接穗大小而定,切口要平整,接穗的长切面向内对准插入母砧切口,注意形成层对准、相互贴紧,用塑料薄膜绑牢封闭严。嫁接后要经常进行检查,发现塑料膜松动要注意补接,加强遮荫防晒,可将剪掉的枝条用作覆盖防晒,注意防治蝽蟓、尺蠖、蚂蚁及卷叶蛾等害虫,可在新梢喷1~2次菊酯类农药,并适时解缚和抹除砧芽。3.3 肥水管理3.3.1 嫁接后管理嫁接后以薄肥勤施为主,定期锄草、松土,使树盘逐年扩大。在第1次新梢老熟后开始施肥,嫁接后第一年在东西向施肥,在树冠两边滴水线处开条沟(宽0.3 m、深0.5 m)进行填埋。翌年在南北向施肥,每次施1.0~1.5 kg有机肥+0.3 kg复合肥,每年结合冬季清园中耕翻土1次,填入绿肥、生石灰等改良土壤。果园采用自然生草或套种绿肥法,在树盘外套种豆科植物、花生等,保留藿香蓟、苜蓿、三叶草等良性杂草,注意保护果园自然天敌和土壤有益生物,只在冬季清园一次。3.3.2结果树管理投产树施肥采用绿色高效施肥方法,即将传统的花前肥、保果肥、壮果肥及采后肥等一年至少4次肥,合并成重施壮果保果肥和施足采后攻梢肥2次肥,并以腐熟农家肥或生物有机肥为主,减少化肥施用量,有利于节本增效,减少农业面源污染。3.3.2.1重施壮果保果肥及时施好壮果肥可补充开花消耗的养分,保证果实需要的营养,减少落果,提高果实品质和产量。施肥时间应选择在疏花后,主要施钾肥,辅以磷肥、氮肥,配施一次5~10 kg腐熟有机肥,并结合喷施1~2次0.3%~ 0.5%磷酸二氢钾。在幼果期至采果前20 d间,喷施杀虫杀菌剂+0.3%~0.5%尿素+0.3%磷酸二氢钾+20 mg/kg赤霉素或防落素,可以更好地起到壮果保果的作用。3.3.2.2 施足采后攻梢肥健壮的结果母枝有助于增加荔枝挂果量[411],采果后施足攻梢肥,可以促秋梢培育优良的结果母枝,为翌年开花结果打好基础。攻梢肥一般在采果后10d内开始施肥,以挂果50 kg计算,有机肥施用量为10 kg,磷肥2 kg,混施3~4 kg复合肥+2 kg尿素,可在施后浇水或在雨后撒施。在末次秋梢中后期施5~10 kg腐熟的花生麸、鸡粪等有机肥,配施钙镁磷肥。3.3.2.3加强土壤水分管理糯米糍荔枝花芽分化需要在冬季(一般在11月中下旬至12月中旬)适度控制水分,在秋梢、花穗抽发期、花蕾期和果实膨大期等要及时适量灌水,特别是要在花穗抽发期保持土壤湿润以促花;遇暴雨或持续降雨时,要及时排涝,以免出现落果、裂果、烂根、树势弱、死亡等问题。3.4 树体管理3.4.1幼树整形修剪未挂果幼树一般轻剪或不剪。分枝较多的可以适当疏剪;枝条太弱的可以适当短截整形,以形成较紧凑、均匀的半圆形或伞形树冠。3.4.2 结果树整形修剪挂果树整形修剪可达到控冠矮化,树高最好控制在3 m以下。在采后10~ 15 d内完成修剪,为防止出现大小年结果现象,修剪方式及程度应根据挂果多少和树体情况来对树冠进行适度回缩,挂果多的要轻剪,适度疏除病枝、弱枝,留下5~7 cm末次秋梢以利于培育健壮的结果枝;挂果少的,做到“六去留平壮”,即从枝条基部剪除病虫枝、枯死枝、荫蔽交叉枝、不完全芽、过密枝、过高直立枝等,保留水平健壮枝,在树冠中央开1.0 m以上的大天窗以培养高光效树形。对结果枝进行短截修剪,基部留3~4片叶子,利于抽发翌年的结果枝,并能使其枝组更新,每年轮换更新1/3~1/4的枝条,留下较强壮的枝条能更好地开花结果。糯米糍荔枝采果后一般要放2次秋梢, 放第一次秋梢有利于恢复树势和培养第二次秋梢,时间在7月下旬至8月上旬,若7月底未发现第一次秋梢有萌动,最好施1次速效化肥促发;第二次秋梢抽发时间一般在9月下旬至10月上旬,该段时间抽发最适合作为翌年结果母枝的秋梢,使得单株结果产量较高[12]。
曾丽兰等糯米糍荔枝在莆田的引种表现及栽培技术- 9 -3.5 防治落果裂果糯米糍荔枝容易发生落果裂果,特别是果实发育后期裂果较严重,可通过以下一系列措施来防治落果裂果。3.5.1 控冬梢促花冬梢老熟时间晚,因营养积累不够而导致开花较晚,其裂果率较高[13]。因此要通过控冬梢促花,保证糯米糍荔枝适时开花,以减少裂果。末次秋梢老熟后,可用40%多效唑600~800倍水溶液喷施叶面进行控梢促花,对已抽发的冬梢可进行人工摘除;对于长势壮旺的荔枝树,宜采用螺旋环割2圈,对树体伤害少愈合快。还可以用旋耕机作断根处理以控冬梢,这样更有利于省工。3.5.2 人工疏花人工疏花可以减少因营养不足导致的落果,提高有效座果率,有效提高产量。时间一般在立春过后,可使用竹鞭敲打花穗,先横扫打断过长花穗,再竖打,减少侧穗及支穗条数,可用主轴抽3~5条侧穗,留2条侧穗,其余摘去。当花穗长至5~8 cm时,用2~6 mL的40%乙烯利+8~10 g的15%多效唑可湿性粉剂兑水15 kg喷洒树冠1~2次。3.5.3 加强水分调控如果幼果期遇高温干燥天气应及时淋水,如果果实膨大期遇长时间雨天应及时排水,可以减少裂果。并结合套种绿肥和地膜覆盖等方式保持土壤湿度,防止土壤干旱,可显著防止裂果。3.5.4 平衡供应所需营养除平衡补充氮、磷、钾等大量元素外,还需及时于果实发育期在地面施钙、钾肥,并喷施硼、铜、锌等叶面肥,但要注意适度施氮肥,过量会造成果实因膨大过快而发生裂果。钙肥宜在冬季果园管理期或幼果期进行施用,可选择贝壳灰作为钙肥,将2~3 kg贝壳灰均匀撒施于成年果树冠下。钾肥宜在果实膨大期施用,应勤施薄施,以防止钾和钙因产生拮抗反应而导致裂果加重,一般每15 d施1次1~2 kg草木灰作为钾肥,撒施于成年果树冠下。在谢花后10 d土施1 kg鸡蛋壳粉和喷施200倍鸡蛋壳粉也能有效防治裂果[14]。3.5.5 使用植物生长调节剂在花蕾期或谢花后及果实着色前用3×10–5萘乙酸喷施,在果实有绿豆大时用10–5乙烯利喷果,间隔1个月再喷一次,辅以环割(一般在10~11月份秋季)、果实套袋、病虫害防控等,保果和防裂果效果更显著。也可在糯米糍荔枝盛花期、果实生长期喷施1 mmol/L GA+0.1 mmol/L Ca(NO3)2,增加原果胶含量以减少裂果[15]。3.6 综合防治病虫害按照“预防为主,综合防治”的植保方针,在科学修剪、生草栽培、清园翻耕等农业防治的基础上,优先采用黄板、太阳能诱虫灯和生物农药等物理、生物防治技术,在化学防治方面尽量少用或不用,要尽量选用低毒高效的农药。病害主要是炭疽病、霜疫霉病等[16]。防治炭疽病在糯米糍荔枝新梢抽生期、花蕾期和幼果期,每隔10 d喷施一次70%甲基托布津800~1 000倍液或75%百菌清;防治霜疫霉病在糯米糍荔枝花蕾期和幼果期,每隔10~15 d喷施一次58%瑞毒霉甲霜锰锌400~600倍液或90%三乙膦酸铝500~600倍液。应重点防治荔枝蒂蛀虫,在糯米糍荔枝谢花后或果实由绿转红时,每隔10~15 d喷施2.5%联苯菊酯2 000~2 500倍液2~3次。桔小实蝇、卷叶蛾、尺蠖、金龟子、毒蛾、木蠹蛾、茶材小蠹、天牛等虫害会严重阻碍糯米糍荔枝健壮秋梢的抽生,从而影响翌年的开花结果,因此要做好杀虫护梢,可在秋梢期施用1~2次菊酯类农药。同时做好冬季清园,刈割杂草并做好树盘覆盖,全园喷施1次0.5%波尔多液、矿物油等, 剪除病虫枝和清除落叶、落地果,并做好无害化处理,减少越冬的病虫害。3.7 适时采收果实糯米糍荔枝应适时采收,有利于克服大小年结果问题。一般在果实八成熟时采收,此时果皮颜色红中带点绿纹,采果时间应选择在上午10点前和下午4点后,以防果实因被太阳暴晒而影响耐贮性,忌雨天和烈日天采收。采取短枝采果法,即采收时用枝剪小心地由果穗基部剪下,不带叶或少带叶,留下枝叶利于发秋梢;采收过程应轻采轻放、轻装轻运,避免果实擦伤,要尽快将采下的果实转移到阴凉处降温,不可将其堆放在地上被日晒雨淋。先采收树势弱和挂果较多的树,有利于恢复树势,促进抽发秋梢,以培育良好的结果母枝。
2025年2月热带农业科学第45卷第2期 10 4 结论糯米糍荔枝能够适应莆田地区的气候条件,易成花结果,与本地主栽品种陈紫相比,果肉更细嫩香甜,不易流汁,内膜褐色更少,焦核率达到90%,莆田的果实成熟期为7月中下旬,较陈紫晚熟约13 d。糯米糍荔枝适宜种植区域较广,20212023年平均产地售价分别为48、42、46元/kg,陈紫荔枝售价分别为32、30、34元/kg,糯米糍经济效益较好。引进推广该品种可以扩大莆田荔枝种植区域,提高莆田荔枝种植面积,调整优化莆田荔枝品种结构,有利于提高经济效益。参考文献[1] 杜朝金, 张汉尧, 罗心平, 等. 分子标记在荔枝和龙眼应用中的研究进展[J]. 中国果树, 2024(1): 1-10+29. [2] 叶翰江, 林锦何, 曾环标. 环剥与环割对糯米糍荔枝增产效应的研究[J]. 中国南方果树, 2014, 43(6): 77-79. [3] 赖永超, 罗诗, 吴美良, 等. 东莞糯米糍荔枝丰产稳产综合栽培技术[J]. 广东农业科学, 2002, 29(4): 29-31. [4] 严婷, 张树飞, 陈庆欣, 等. “糯米糍” 荔枝栽培生理与技术研究进展[J]. 中国南方果树, 2022, 51(5): 200-206. [5] 莆田市统计局, 国家统计局莆田调查队. 莆田统计年鉴(2023)[M]. 北京: 中国统计出版社, 2023. [6] 中华人民共和国农业部植物新品种特异性、一致性和稳定性测试指南荔枝:NY/T2564—2014[S]. 北京: 中国标准出版社, 2014. [7] 欧良喜, 陈洁珍. 荔枝种质资源描述规范和数据标准[M]. 北京: 中国农业出版社, 2006. [8] 田健伟. 糯米糍荔枝抗逆高效栽培技术[J]. 柑桔与亚热带果树信息, 2003, 19(2): 32-33. [9] 陶忠良, 高爱平, 周兆德. 气象条件对荔枝产量的影响研究综述[J]. 中国南方果树, 2001, 30(4): 29-31. [10] 何湖清, 吴雅芳. 莆田市气候特点及气温变化特征分析[J]. 农业灾害研究, 2020, 10(4): 60-61+101. [11] 倪耀源, 吴素芬. 荔枝栽培[M]. 北京: 农业出版社, 1990. [12] 李建国, 陈华. 糯米糍荔枝末次秋梢抽生期对花期、花质及着果的影响[J]. 中国南方果树, 1998, 27(4): 24-25. [13] 杨惟智, 郑杰, 陈庆芳, 等. 糯米糍荔枝的裂果规律研究与预防措施[J]. 上海蔬菜, 2022(6): 59-60+64. [14] 麦达文. 鸡蛋壳粉对糯米糍荔枝的防裂果试验[J]. 广东农业科学, 2006, 33(2): 35-36. [15] 彭坚, 席嘉宾, 唐旭东, 等. 叶面喷施Ca(NO3)2和GA对‘糯米糍’荔枝裂果的影响[J]. 园艺学报, 2001, 28(4): 348-350. [16] 康丽雪. 糯米糍荔枝优质丰产栽培技术[J]. 福建果树, 2010(3): 55-56. (责任编辑林海妹)
2025年2月热带农业科学第45卷第2期Feb. 2025 CHINESE JOURNAL OF TROPICAL AGRICULTURE Vol.45, No.2 收稿日期 2024-03-04;修回日期 2024-04-10 基金项目 海南特有灭绝级药用植物爪耳木的繁育体系构建与自然回归试验研究(No. 321MS090);农业野生植物资源等调查(No. 13230203);2024年国家热带植物种质资源库。第一作者汤欢(1986—),男,博士,助理研究员,研究方向为植物种质资源收集保存鉴定,E-mail:tanghuan_a@163.com。通信作者 王清隆(1983—),男,硕士,副研究员,研究方向为植物资源开发利用与植物分类,E-mail:qlwang1983@163.com。野外灭绝级植物爪耳木的繁育汤欢羊青王茂媛王祝年刘咲頔王清隆(中国热带农业科学院热带作物品种资源研究所/海南热带植物园/农业农村部中药材生物学与栽培重点试验室/农业农村部热带农业野生植物基因资源鉴定评价中心/ 海南省热带药用植物工程研究中心海南海口 571101)摘要爪耳木作为国家二级保护珍稀濒危物种,为我国特有植物,目前在IUCN上仍被列为灭绝级(EX)植物。爪耳木在2013年被科研人员再次发现后,科研人员围绕爪耳木的资源调查以及生物学特性、种苗繁育等方面开展了相关研究。爪耳木目前作为一种在野外已灭绝的植物,对其开展相关研究和重新回归野外具有重要价值,对爪耳木属植物系统发育和进化的研究也有重要参考价值。文章对爪耳木开展繁育经验总结发现,爪耳木可以通过种子繁殖、叶和根扦插、根蘖繁殖获得新植株。爪耳木的成功大量繁育将为爪耳木后期的野外回归做好准备,并有助于其野外种群的恢复和保护以及濒危机制探讨等一系列研究的深入开展。关键词爪耳木;灭绝级;扦插;根蘖;野外回归中图分类号S533 文献标识码 A DOI: 10.12008/j.issn.1009-2196.2025.02.003 Research on the Propagation of Lepisanthes unilocularis, An Extinct Plant in the Wild TANG Huan YANG Qing WANG Maoyuan WANG Zhunian LIU Xiaodi WANG Qinglong (Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences/Hainan Tropical Botanical Garden/Key Laboratory of Biology and Cultivation of Herb Medicine, Ministry of Agriculture and Rural Affairs, P. R. China/Identification and Evaluation Center of Tropical Agricultural Wild Plant Gene Resources, Ministry of Agriculture and Rural Affairs, P. R. China / Hainan Provincial Engineering Research Center for Tropical Medicinal Plants, Haikou 571101, China) Abstract Lepisanthes unilocularis, a rare and endangered species protected at the national level with second class, is the Chinese endemic plant. It is still listed as an extinct plant by the International Union for Conservation of Nature. Lepisanthes unilocularis was rediscovered by researchers in 2013. Researchers have carried out relevant research on resource investigations, biological characteristics, seedling propagation, etc. Lepisanthes unilocularis is now an extinct plant in nature. It is of great value to carry out relevant research and reintroduction into the wild. It also has vital reference value for the study of the phylogeny and evolution of Lepisanthes. In this paper, the propagation experience summary of Lepisanthes uniloculariswas discussed. The results revealed that new plants could be obtained via seed propagation, leaf and root cuttings, and root sprout propagation. Successful mass propagation will prepare for the later reintroduction of Lepisanthes unilocularis. And facilitate the in-depth development of a series of related studies, including the restoration and protection of the wild population and the mechanism of its endangerment. KeywordsLepisanthes unilocularis; extinct; cutting; root sprout; reintroduction 爪耳木Lepisanthes unilocularis Leenhouts为无患子科鳞花木属常绿灌木,为我国特有植物,特产海南[1-3],目前在IUCN上被列为灭绝级(EX)植物[4]。爪耳木1935年被发现于海南乐东佛罗,迄今已半个多世纪没有再次被采到,是一种可能或接近于灭绝的植物。爪耳木属是典型的热带亚洲属,分布中心在加里曼丹岛,我国仅海南有1种,对研究和论证海南植物区系与上述地区的关系有一定意义。爪耳木又是海南热带滨海砂生植被中为数甚少的树种之一,极能耐旱耐脊,应是
2025年2月热带农业科学第45卷第2期12 这类地区较理想的造林树种[5]。爪耳木在新发布的《国家重点保护野生植物名录》中被列为二级保护植物[6]。1935年,中国著名植物采集家刘心祈先生在海南崖县佛罗(今乐东佛罗镇)附近沿海疏林间发现并采到1份带果的爪耳木标本(S. K. Lau 5773),多年之后人们才根据此份带果的爪耳木标本将爪耳木正式发表出来[7],随后很长一段时间爪耳木都未被科研人员发现。直到2013年,本团队科研人员在乐东尖峰镇再次发现了爪耳木野外居群,在此之前,人们一度认为爪耳木已经灭绝。爪耳木再次被发现至今已有10年,这期间,科研人员围绕爪耳木野外居群分布和环境因子等资源调查[8-9]、爪耳木的叶片功能性状[10]、濒危机制与生物信息学挖掘[11]、染色体倍性与基因组大小测定[12]、包含爪耳木花和果特征的物种中文和英文形态描述[8-9]、生物学特性、人工繁育、开花物候、种子萌发特性以及基因组测序等方面进行了较系统的研究工作。以此为基础,本研究以成功引种并扩繁的野外灭绝级植物爪耳木为研究对象,采用种子繁殖、根蘖繁殖、扦插、高空压条、组织培养等方法,通过不断优化试验,寻找爪耳木可行的扩繁途径,及时进行爪耳木繁育经验总结。开展爪耳木繁育技术探讨及经验总结,旨在得到一些切实可行的爪耳木繁育技术,大量繁育爪耳木种苗,为爪耳木后期的野外回归做好准备,并有助于其野外种群的恢复和保护以及濒危机制探讨等一系列研究的深入开展。1爪耳木繁育技术爪耳木在2013年被科研人员再次发现后,引种了5株到海南儋州的农业农村部儋州热带药用植物种质资源圃。通过近十年的努力,目前已成功繁育出3 000多株爪耳木种苗。以下为本团队摸索出的成功繁育爪耳木的一些方法,包括使用种子繁殖、复叶和根扦插繁殖,以及根蘖繁殖。1.1种子繁殖爪耳木为单性花,雌雄同株,盛花期为1月中旬,以雌花开放为主;雄花一般到了3月底,天气比较干旱时才有少量开放;整个花期雄花较雌花明显偏少,雌雄花比例高达3 000∶1,高温干旱有助于雄花开放,但量极少,若遇上连续低温或者下雨就极不容易开雄花,且单朵雄花的花期仅约1 d时间。以上原因致使爪耳木整个居群的花粉量少、雌雄花期不遇,而影响传粉和受精,加上多种因素引起的种胚败育,以及一些动物咬食果实,导致最终成熟的爪耳木果实极少,进一步加剧其濒危。人工授粉可以显著提高爪耳木坐果率和结实率,人工授粉结果率可高达85%[11]。因此,可以人工辅助授粉,以增加其结实率。盆栽3年以上达到开花期的种苗,通过人为控水,保持高温干旱条件,促进爪耳木雄花在盛花期大量开放,以帮助爪耳木完成授粉。此外,通过套袋并及时采收,可增加收获的种子量。目前,每年都能从引种扩繁的爪耳木上采收到一批成熟种子(图1),检测其发芽率基本都能正常发芽(图2)。图1从引种扩繁的爪耳木上采收的成熟种子图2爪耳木种子发芽率检测本团队研究的爪耳木种子萌发试验结果为:种子共播53粒,种子随采随播于富含有机质的沙壤土中,4 d后开始发芽,12 d后芽开始露土,共计种子发芽数为33粒,发芽率约为62.3%。爪耳木种子不耐低温,经过3℃低温储藏处理1周后,发芽率仅为13%。种子生活力试验结果表明:爪耳木的种子为顽拗性种子,只要种子失水超过
汤欢等野外灭绝级植物爪耳木的繁育-13 -30%就会失活。由于爪耳木果实成熟期多为3月底至4月初左右,海南的气温白天较高,可达37℃,而且高温干旱时间较长,一般来说爪耳木种子在自然条件下第3 d以后就会失活[11]。鉴于此,在爪耳木开花期人工辅助授粉,并及时采摘成熟的种子,即采即播,以增加其发芽率。1.2根蘖繁殖爪耳木虽然在有性生殖上存在一定障碍,但其在无性繁殖上有补尝机制,即发生根蘖——根蘖更新是无性繁殖的一种方式,母株通过横走侧根,在其周边2 m左右内长出不定芽,不定芽产生无性繁殖体。这些繁殖体定居成功后就发展为潜在的独立植株,即无性系子株,从而达到种群更新的目的。由于不定芽产生的无性系子株在一段时间内仍与母株根系相连接,各繁殖体间可通过物质交换传输的生理整合作用共同利用生境内的资源,与有性繁殖相比,大大降低了生长代价[8, 13-16]。利用爪耳木根蘖能力强的特点,本团队开展了爪耳木的根蘖繁殖试验。结果发现,利用盆栽控水处理可快速诱导爪耳木的根产生大量不定芽(图3、4)。故可利用此方法处理获得大量的爪耳木根蘖苗进行快速根蘖扩繁。此外,鉴于一些学者研究发现实施人工切断水平根的方式有利于促进根蘖苗的萌发[17-19],故还可时常进行断根处理以刺激爪耳木萌发更多根蘖苗。1.3扦插繁殖基于相关文献资料[20-26],制定扦插方案,并于2023年1月前后再次开展了爪耳木扦插繁殖试验,以下为此次爪耳木扦插试验情况。1.3.1试验地概况试验地位于海南省儋州市农业农村部儋州热带药用植物种质资源圃(以下简称“南药圃”),所在地经纬度为19°30′44″N,109°30′20″E,属于热带海洋性季风气候,四季温暖。1.3.2试验材料试验材料来自南药圃多年生爪图3盆栽控水栽培处理爪耳木的根萌出芽点情况图4盆栽控水栽培处理爪耳木的根产生大量不定芽耳木母树。扦插插穗采自母树健壮无病虫害的叶片、茎干和根。叶片取自地栽爪耳木植株中上部较健壮肥硕的整个奇数羽状复叶,采摘时尽量将整个叶柄甚至带一点枝上的皮一起取下,以保证整个复叶的完整,取完叶片后用剪刀剪去部分小叶;茎干取自地栽爪耳木植株中新抽发出的较肥硕且近木质化枝条上的茎段,将其基部斜剪,制作成长度约10 cm的插穗;根取自5盆盆栽的爪耳木,先将盆横放,轻轻摇松,再慢慢从盆中横向拔出爪耳木,抖掉部分泥土,剪出其中直径在0.5 cm以上的根,再剪取长度在5 cm以上的带芽点的根段作为插穗。1.3.3试验方法1.3.3.1试验前准备扦插前使用浓度0.5%左右的高锰酸钾溶液对基质进行消毒。在装有已消毒蛭石基质的带盖扦插育苗盒中,洒入适量清水,使基质手握成团松手即散即可,再用剪刀或硬棒一一打洞,在洞内放入处理好的插穗后再轻轻压实,使插穗保持直立不倾倒状态,再盖上扦插盆自带的透明塑料盖即可。
2025年2月热带农业科学第45卷第2期14 1.3.3.2试验设计爪耳木扦插研究分别设置叶片、茎干和根这3种插穗,不同浓度的IAA(生长素)、IBA(吲哚丁酸)和NAA(α萘乙酸)生长调节剂配比和不同浸泡时间开展扦插试验,研究不同插穗、不同生长调节剂和不同浸泡时间对爪耳木扦插的影响,具体处理如下。将爪耳木的叶分别于50、100、200、400、800 mg/L的IAA、IBA、NAA以及清水中浸泡30、60、120 min,其叶再分别于200、400、800 mg/L的IAA+IBA、IAA+NAA、IBA+NAA、IAA+IBA+ NAA中浸泡30、60、120 min;将爪耳木的茎于800 mg/L的IAA、IBA、NAA中浸泡120 min;将爪耳木的根分别于50、100、200、400、800 mg/L的IAA、IBA、NAA以及清水中浸泡120 min。用高锰酸钾消毒后的蛭石为扦插基质进行扦插,每个处理10个插穗,约90 d后统计生根率等。1.3.4试验结果近年来,本团队多次开展爪耳木扦插试验,但对爪耳木所做的所有试验,其茎干和枝条扦插都以失败告终。本团队对爪耳木茎干用常规扦插方法暂时还无法获得成活的扦插苗,对其叶片用常规扦插方法加上保温保湿可以大量获得扦插苗(图5);其根因具有根蘖特性,剪成长度在5 cm以上的带芽点的根段作为插穗基本都能长成正常的扦插苗(图6)。此外,发现在800 mg/L的IBA中浸泡120 min再扦插,其扦插成苗率较高。2野外回归与保护在科研人员精心引种繁育下,运用种子繁殖、扦插、根蘖繁殖等繁殖方式,目前已成功繁育出3 000多株爪耳木种苗。爪耳木濒危状态已基本解除,暂无灭绝之虑。鉴于爪耳木原生境为乐东黎图5爪耳木根扦插后长出植株图6爪耳木叶片扦插后长出根族自治县佛罗镇和尖峰镇附近滨海沙地上的疏林灌丛,本团队将会从已繁育的3 000多株爪耳木中挑选出一部分较健壮的植株,分步骤开展回归试验。前期计划先联系其原生境周边农户,先引种到农户家自留地,待成功引种后,再进一步从农户家自留地移栽到附近适宜的滨海沙地上的疏林灌丛中,同时在收获种子后也在农户家自留地及附近滨海沙地上的疏林灌丛中开展原生境种子繁殖试验,使爪耳木能尽快成功回归野外,并在其回归野外后加强对当地居民的宣传工作,减少人为破坏。此外,后期还将进一步让爪耳木进入种子库、离体库、DNA库、植物园等进行保存,为爪耳木的保护提供更多的安全保障。3讨论与结论野外灭绝级植物爪耳木,由科研人员在近80年后再次发现并引种扩繁后,现已繁育出3 000多株种苗,基本避免了灭绝风险。本团队通过近10年来始终不懈地开展爪耳木繁育研究,发现爪耳木可以进行种子繁殖、使用其叶片和根进行扦插繁殖以及通过根蘖繁殖而获得大量种苗。在热带地区顽拗性种子占比非常大[27]。爪耳木种子为顽拗性种子,不耐脱水和低温,贮藏难度较大。在爪耳木种子繁殖试验中发现,爪耳木虽然结果量极少,且果实落地后超过3 d就基本失活,但可以通过人工辅助授粉增加其结果量,并及时采摘成熟果实,即采即播,增大爪耳木等顽拗性种子的萌发率,加速扩繁大量植株。爪耳木种子随采随播,是保证其大量发芽的关键。萌蘖在植物界中普遍存在,是植物的一种重要繁殖方式,特别是在木本植物中,萌孽是维持树木更新的重要方式之一[28]。一些研究者认为,森林植物的萌蘖能力既受生境、干扰、资源水平等外在
汤欢等野外灭绝级植物爪耳木的繁育-15 -因素的影响,又受内部营养水平和激素合成等控制,是两者共同作用的结果[29]。根据本团队多年的野外实地观察以及相关文献资料[8, 13-19, 28, 30, 31]查阅发现,许多植物都容易发生根蘖,如构树、刺槐、枣、杨属中一些物种(胡杨、毛白杨等)、大青属中一些物种(大青、赪桐、烟火树等)、菊科中一些物种(紫茎泽兰、艾纳香、五月艾、艾等)、芭蕉属中一些物种、姜科中一些物种(红茴砂、茴香砂仁、砂仁等)、无患子科中一些物种(爪耳木、文冠果、细子龙等)、禾本科中一些物种、接骨草、接骨木、火炬树、火焰树、木荷、海南巴豆、海南椴、香果树、杜仲、泡桐、树胡椒、银荆、银杏、香椿、臭椿、沙棘、女贞、通脱木、吴茱萸等,这些植物都具有较强的根蘖性。爪耳木虽然在有性生殖上存在一定障碍,但其在无性繁殖上有补尝机制,即像上述许多植物一样容易发生根蘖。在爪耳木根蘖繁殖试验中发现,利用盆栽控水处理可快速诱导根产生大量不定芽。利用断根等处理也能较好地刺激爪耳木萌发出更多的根蘖苗。因此,在实际生产中可以充分利用爪耳木极强的根蘖性大量繁育爪耳木种苗。有学者研究发现,根原基类型、插穗的生根类型、茎的解剖结构是决定扦插难易程度的三大重要影响因素[20, 32-33]。关于野外灭绝级植物爪耳木扦插方面的研究,除本团队有开展外,暂未获知有其他学者开展。爪耳木通过种子培育成苗周期长,而通过扦插繁殖、根蘖繁殖等无性繁殖方式培育种苗可极大缩短育苗周期。故本团队近年来多次开展爪耳木扦插繁殖试验,旨在通过探索不同扦插插穗和生长调节剂配比对爪耳木扦插的影响,探究爪耳木适宜的扦插条件,以期快速繁育爪耳木种苗。之前,本团队开展的爪耳木扦插繁殖试验结果表明,爪耳木茎干为极难扦插生根类型,采用常规的生根剂(ABT、IBA、NAA、国光生根粉等)不同浓度不同时间处理,都不能诱导生根,而通过清水清洗24 h后再处理和通过茎段环剥处理等长出愈伤组织再用生根剂处理也都不能诱导生根,说明爪耳木在茎干扦插繁殖上扩繁难度较大[11]。但对于爪耳木的叶片,在利用带盖扦插育苗盒以保证其空气相对湿度接近于饱和状态下,能够顺利得到其叶片扦插苗,但在之前的试验中未用育苗盒,空气湿度不高则基本不能获得叶片扦插苗,由此可知,空气湿度是影响爪耳木叶片扦插成活的关键因素。爪耳木的根因其极强的根蘖特性,极易在根上萌发出不定芽,故利用其带不定芽的根进行扦插相对比较容易产生扦插苗。在近10年间对爪耳木开展的扦插试验中,用茎干都不成功,通过不断思考改进,试着用其复叶进行扦插并最终成功获得了植株,之前笔者仅发现在一些秋海棠科、爵床科中的一些种类,还有多肉类植物中,能通过叶片扦插获得新植株,但这些植物多是一些草本类植物,且是单叶,从没想像过像爪耳木这类灌木,其茎干目前都未能成功进行扦插获得新植株,其叶片,而且是复叶,却可以通过扦插获得新植株,目前推测这可能是因为其复叶膨大的叶柄基部聚集有较丰富的促进生长的营养物质,使其能在扦插后长出根和芽,此发现也可作为其他具有膨大叶柄类植物扩繁时考虑的一个可能的途径。扦插时,将带完整膨大叶柄的复叶基部倾斜插入透气透水的基质中再稍稍压紧,在带盖扦插育苗盒空气相对湿度接近于饱和状态下培养,其叶片扦插成苗率能达到85%以上。此外,本团队也使用爪耳木的花药、花粉、种子、嫩芽进行多次的组织培养,并开展高空压条试验,但多年来的相关试验均未能成功获得植株,其原因还有待更深入系统的相关研究。通过上述方法培育获得大量爪耳木植株后,再通过一步步的野外回归试验,有望实现爪耳木野外种群的逐步恢复。鉴于爪耳木目前在IUCN上仍被列为灭绝级(EX)植物,期盼相关人员能对爪耳木的濒危状况进行重新评估,及时更新其濒危等级为野外灭绝级(Extinct in the Wild,EW),并加大宣传和保护力度,以期让更多的人能够参与到爪耳木的保护和研究工作中。参考文献[1]中国科学院中国植物志编辑委员会. 中国植物志: 第四十七卷第一分册[M]. 北京: 科学出版社, 1985: 26-27. [2]Wu Z Y, Raven P H. Flora of China: Volume 12[M]. Beijing: Science Press, 2007: 15. [3]广东省植物研究所. 海南植物志: 第三卷[M]. 北京: 科学出版社, 1974: 82.
2025年2月热带农业科学第45卷第2期16 [4]IUCN 2023. The IUCN Red List of Threatened Species. Otophora unilocularis. [EB/OL]. Version 2022-2. [2023-08- 09]. https://www.iucnredlist.org/search?query=Otophora%20 unilocularis&searchType=species. [5]傅立国. 中国植物红皮书稀有濒危植物: 第一册[M]. 北京: 科学出版社, 1991: 594. [6]国家林业和草原局, 农业农村部. 国家重点保护野生植物名录(2021年第15号)[EB/OL]. (2021-09-07) [2023-08-09]. https://www.gov.cn/zhengce/2021-09/07/content_5727413.htm. [7]Leenhouts P W. A revision of Lepisanthes (Sapindaceae) [J]. Blumea, 1969, 17(1): 73-75. [8]王清隆, 羊青, 王茂媛, 等. 海南特有灭绝级(EX)植物爪耳木的资源调查[J]. 热带农业科学, 2018, 38(2): 56-60. [9]Wang Q L, Tang H, Wang M Y, et al. The rediscovery of Lepisanthes unilocularis (subgen. Otophora; Sapindaceae), a species endemic to Hainan, China [J]. Nordic Journal of Botany, 2018, 36(4): 1-4. [10]刘咲頔, 王祝年, 王清隆, 等. 海南特有灭绝级植物爪耳木的叶片功能性状[J]. 贵州农业科学, 2020, 48(2): 8-11. [11]刘咲頔. 灭绝级植物爪耳木濒危机制与生物信息学挖掘鉴定爪耳木中GA相关基因研究[D]. 海口: 海南大学, 2020. [12]李英英, 刘咲頔, 王清隆, 等. 基于流式细胞术的爪耳木染色体倍性与基因组大小测定[J]. 现代农业科技, 2022 (18): 84-86+94. [13]田登娟, 聂恺宏, 张增悦, 等. 中国沙棘根蘖能力及非结构性碳水化合物对平茬高度的响应[J]. 西北农林科技大学学报: 自然科学版, 2023, 51(9): 70-83. [14]武逢平, 李俊清, 李景文, 等. 胡杨(Populus euphratica)在额济纳绿洲三种生境内的根蘖繁殖特性[J]. 生态学报, 2008, 28(10): 4 703-4 709. [15]刘杰, 孙宇涵, 袁存权, 等. 林木根萌复幼方式的研究进展[J]. 分子植物育种, 2022, 20(14): 4 867-4 872. [16]马亚平, 白琳云, 徐伟荣, 等. 灵武长枣嫁接和根蘖繁殖植株果实转录组差异分析[J]. 分子植物育种, 2021, 19(16): 5 297-5 306. [17]王子康, 焦阿永, 凌红波, 等. 不同灌溉模式下胡杨断根处理根蘖繁殖特征[J]. 干旱区研究, 2022, 39(4): 1 133- 1 142. [18]郭连金, 薛苹苹, 邵兴华, 等. 香果树根萌苗生长特性及影响因子分析[J]. 植物科学学报, 2015, 33(2): 165-175. [19]田永祯, 司建华, 程业森, 等. 荒漠河谷胡杨残林复壮更新试验研究[J]. 干旱区资源与环境, 2009, 23(9): 155-159. [20]孙雪莲, 杨楚童, 胡亚楠, 等. 植物扦插生根机理的研究进展[J]. 农学学报, 2021, 11(10): 33-40. [21]贾志远, 葛晓敏, 唐罗忠. 木本植物扦插繁殖及其影响因素[J]. 世界林业研究, 2015, 28(2): 36-41. [22]李焕勇, 刘涛, 张华新, 等. 植物扦插生根机理研究进展[J]. 世界林业研究, 2014, 27(1): 23-28. [23]伏桂仙, 季玉松, 陶俊. 植物扦插繁殖影响因素综述[J]. 分子植物育种, 2023, 21(6): 2 080-2 088. [24]潘向艳. 火焰芦莉扦插繁殖影响因素研究[J]. 热带农业科学, 2023, 43(12): 23-28. [25]邱才飞, 邱小英, 陈金, 等. NAA和IBA浸沾浓度及时间对地菍插枝发根的影响[J]. 热带农业科学, 2023, 43(3): 47-50. [26]邓丽丽, 秦惠珍, 熊忠臣, 等. 药食两用植物五指毛桃扦插技术研究[J]. 热带农业科学, 2022, 42(2): 23-30. [27]Subbiah A, Ramdhani S, Pammenter N W, et al. Towards understanding the incidence and evolutionary history of seed recalcitrance: An analytical review[J]. Perspectives in Plant Ecology, Evolution and Systematics, 2019, 37: 11-19. [28]冯辉, 王海洋, 张楠, 等. 森林木本植物萌蘖更新的研究进展[J]. 贵州农业科学, 2011, 39(9): 177-180+184. [29]Zhu W Z, Wang J X, Luo C R, et al. Progresses of studies on forest sprout regeneration[J]. Sclentla Silvae Sinicae, 2007, 43: 74-82. [30]刘常幸, 金毅, 余建平, 等. 古田山茶湾样地甜槠木荷林根萌特征分析[J]. 浙江大学学报: 理学版, 2014, 41(5): 573-582+592. [31]康向阳. 杜仲良种选育研究现状及展望[J]. 北京林业大学学报, 2017, 39(3): 1-6. [32]Hatzilazarou S P, Syros T D, Yupsanis T A, et al. Peroxidases, lignin and anatomy during in vitro and ex vitro rooting of gardenia (Gardenia jasminoides Ellis) microshoots[J]. Journal of Plant Physiology, 2006, 163(8): 827-836. [33]Zhang W, Fan J, Tan Q, et al. The effects of exogenous hormones on rooting process and the activities of key enzymes of Malus hupehensis stem cuttings[J]. PLoS One, 2017, 12(2): e0 172 320. (责任编辑龙娅丽)
2025年2月热带农业科学第45卷第2期Feb. 2025 CHINESE JOURNAL OF TROPICAL AGRICULTURE Vol.45, No.2 收稿日期 2024-01-08;修回日期 2024-05-06 基金项目 国家茶叶产业技术体系(No. CARS-19)。第一作者刘春艳(1982—),女,硕士,讲师,研究方向为茶文化及茶树生理生态,E-mail:215849932@qq.com。通信作者 赵远艳(1982—),女,硕士,高级农艺师,研究方向为茶树栽培管理,E-mail:ynkmndzhao@163.com。不同调酸措施对酸化茶园土壤改良以及茶叶产量的影响刘春艳1赵远艳2焦婷婷1,3文勤枢4胡余花1熊昌云1,3(1. 云南农业大学茶学院云南昆明 650201;2. 普洱市茶叶科学研究所云南普洱 665000;3. 云南农业大学普洱校区茶叶研究所云南普洱 665000;4. 云南德宏州茶叶技术推广站云南德宏 678400)摘要为进一步探索酸化土壤改良技术的最佳效果及方法,根据茶树良种场茶山分场茶园土壤酸性状况,制定酸化土壤改良对比试验方案,连续施用2年,并对试验茶园的土壤pH以及茶叶产量进行测定。结果表明,不同调酸措施均能够提高茶园土壤pH,其中,处理A、B的pH增加最显著,与处理前相比,分别增加了2.13和2.22个pH单位,且不同处理间的产量也存在一定差异;与CK相比,处理E增产率最高,处理1年后的增产率为9.27%,处理2年后的增产率为11.44%。综上可知,双飞粉、橡胶籽油枯、钙镁磷、尿素、阿姆斯微生物菌肥均具有调节土壤酸化的作用,双飞粉会使土壤pH在短时间内快速上升,橡胶籽油枯、钙镁磷、尿素配合施用在提高土壤pH的同时,也能在一定程度上提高茶叶产量。关键词酸化茶园土壤;酸度;茶叶产量;改良效果中图分类号S533 文献标识码 A DOI: 10.12008/j.issn.1009-2196.2025.02.004 Effects of Different Acid-Regulating Measures on Soil Improvement and Tea Yield in Acidified Tea Plantations LIU Chunyan1 ZHAO Yuanyan2 JIAO Tingting1,3 WEN Qinshu4HU Yuhua1 XIONG Changyun1,3(1. College of Tea, Yunnan Agricultural University, Kunming, Yunnan 650201, China; 2. Pu’er Tea Scientific Research Institute, Pu’er, Yunnan 665000, China; 3.Tea Research Institute, Yunnan Agricultural University , Pu’er, Yunnan 665000, China; 4. Yunnan Dehong Tea Technology Promotion Station, Dehong, Yunnan 678400, China) Abstract To further explore the best method for improving acidified soil, this study developed a comparative trial program for improving acidified soil based on the acidic conditions of tea plantation soil in the Tea Mountain Branch of the Tea Tree Breeding Farm, which was applied continuously for two years, and the soil pH value and tea yield of the test tea plantation were measured. The test results revealed that different acid-regulating measures improved the soil pH value of tea plantations,among which the pH values of treatment A and treatment B increased the most significantly. Compared with that before treatment, the pH value increased by 2.13 and 2.22 pH units, respectively, and there were also some differences in yield between different treatments. Compared with CK, treatment E had the highest yield increase rate, the yield increase rate at treatment 1year was 9.27%, and the yield increase rate at treatment 2 years. In summary, double fly powder, rubber seed oil cake, calcium and magnesium phosphorus, urea, and Ams microbial fertilizer play roles in regulating soil acidification application, and dolomite powder causes the soil pH value to increase rapidly for a short period of time. Rubber seed oil wilt, calcium and magnesium phosphorus, urea, and the application of these fertilizers improve soil pH at the same time, but also, to a certain degree, increase the yield of tea leaves. Keywords acidified tea plantation soil; acidity; tea yield; improvement effect 茶树是多年生植物。作为我国重要的经济作物之一,由于其具有较好的经济效益,近年来茶树种植面积不断扩大,土壤酸化问题也日益突出[1-2]。茶树是喜偏酸性植物,最适宜生长的土壤pH在4.5~5.5,但土壤过酸会使土壤中盐基性阳离子减少,影响茶树生长和养分的吸收,从而影响茶叶
2025年2月热带农业科学第45卷第2期18 产量及品质,成为制约茶产业发展的一个重要因素。Shiyu等[3]对云南省不同种植年限的茶园土壤pH进行调查研究发现,0~40 cm土层pH呈增高趋势,40~80 cm土层pH无明显变化,80~100和100~120 cm土层pH呈酸下降趋势,认为有机肥不能缓解深层土壤的酸化现象。据统计,我国茶园土壤平均pH为4.73,41%的茶园土壤pH处于4.5~5.5,52%的茶园土壤pH在4.5以下[4]。茶园土壤酸化不仅会导致土壤理化性质变差、改变土壤结构、降低土壤的供肥能力和光合作用,也会导致茶园土壤的其他生态环境问题[5-6]。因此,及时采取调酸措施缓解茶园土壤酸化问题,防止茶园土壤进一步退化是目前亟需解决的问题。现阶段茶园土壤酸化修复的措施主要有化学改良[7]、生物改良[8]、有机改良[9]、新型改良剂[10]、合理施肥[4]。双飞粉属于含有碳酸钙等石灰类物质,施入土壤后,能中和土壤酸性,提升土壤pH,但长期施用碳酸钙类土壤改良剂会出现土壤板结、阳离子不平衡、有机质含量下降等不良影响[11];大量研究表明,配施有机肥是稳定和提高土壤pH的重要途径,可以增强土壤抗酸化能力和缓冲性能,改善土壤性状[12-15]。厩肥或高温堆肥等有机肥碱度较高,所以使用厩肥或高温堆肥能够有效控制农田土壤酸化,同时也能节约成本,提高资源利用率[16],钙镁磷肥作为一种碱性矿质肥料,能够提供植物所需的磷、钙、镁、硅等元素[17]。有研究表明,施用钙镁磷肥能显著提高土壤pH,改良酸性土壤,并能通过降低土壤重金属活性,提升作物营养与品质[18-19]。微生物菌肥能够提高土壤内有益细菌的含量和活力、抑制有害细菌的生长、降低土传病害的发生率,增产提质,提高农作物的抗病性[20]。目前,对于改良剂与有机肥混合的方式研究较少,因此本研究针对茶园土壤酸化问题,探究双飞粉、橡胶籽油枯、钙镁磷、尿素、阿姆斯微生物菌肥5种改良剂配合施用对茶园土壤酸碱度以及茶叶产量的影响,分析不同调酸措施的改良效果,以期寻找合理的施用技术。1材料与方法1.1材料1.1.1试验地概况试验茶园位于云南省普洱茶树良种场茶山分场示范茶园(云南省普洱市),地理坐标为东经100°51′43,北纬22°44′35,海拔1 325 m。1.1.2试材茶树品种为云抗10号,树龄22年,茶树长势好、年干茶产量132.4 kg/亩,土壤为红壤。1.2方法1.2.1 试验设计试验共设计5个处理,各处理进行随机排列,每个处理设3个重复,每个重复长度为80 m,详见表1。于2020年11月5日,结合冬季深耕施肥,按配方要求将试验材料施入施肥沟(深宽度30 cm×30 cm),并盖好施肥沟,要求土块打碎平整。第二次施肥时间为2021年11月24日,本次施肥取消双飞粉的使用,土壤改良期间记录各处理的鲜叶产量。表1不同处理土壤改良方法单位:kg/亩处理编号双飞粉(CaCO3)橡胶籽油枯钙镁磷尿素阿姆斯微生物菌肥CK A 261 B 261 2 C 261 150 25 10 D 150 25 10 2 E 150 25 10 注:1亩≈667 m2。1.2.2项目测定(1)茶叶产量测定:调查茶芽密度和百芽重,每处理随机抽样3点,每点调查0.09 m2(30 cm× 30 cm),记录各调查区内的茶芽数,最后折算成每平方米的茶芽数。从调查区内随机采摘3组茶芽,每组100个称重记载。产量:各小区每次采摘后单独称量芽头重量(一芽二叶),最后各批次重量相加即为各小区的总产量。(2)茶园土壤pH测定:采用 pH 计测定。1.2.3数据分析运用 Excel 2021、SPSS 27.0 和Origin 2022软件对数据进行处理分析,其中,
刘春艳等不同调酸措施对酸化茶园土壤改良以及茶叶产量的影响-19 -Excel 2021进行数据统计,SPSS 27.0软件用于单因素方差分析、显著性分析,Origin 2022进行作图。2结果与分析2.1调酸措施对茶园土壤pH的影响由图1可知,通过不同的土壤酸化改良措施,茶园土壤的pH呈现不同程度的变化。处理1年后,0~20 cm土层处理pH除处理D较CK下降了0.02个单位,与CK无显著性差异外(图1-A),处理A、B、C、E依次显著提高了1.24、1.33、0.73、0.10个单位。处理2年后,与CK相比,处理A、B、C分别显著提高了1.21、0.32、0.21个单位,处理D、E分别降低了0.05、0.16个单位,处理D与CK无显著性差异,处理E差异显著下降。20~40 cm土层处理1年后与CK相比,处理A、B、C的pH分别显著提高了0.22、0.27、0.11个单位,处理D、E分别显著降低了0.14、0.1个单位,处理2年后与CK相比,处理A、B、C分别提高0.17、0.03、0.25个单位,处理A、C具有显著上升趋势,处理B无显著性差异,处理D、E分别降低了0.03、0.21个单位,处理D无显著性差异,处理E显著下降(图1-B)。不同小写字母表示差异显著(p<0.05)。下同。图1土壤改良措施对土壤pH的影响2.2调酸措施对芽头密度的影响由表2可知,不同季节芽头密度差异不大,春季、夏季和秋季的芽头密度变异系数依次为5.50%、10.50%、12.77%。与对照(CK)相比,春季处理A、处理B、处理C、处理D的芽头密度均有显著性差异(p<0.05),处理E无显著性差异(p>0.05),所有处理芽头密度均高于CK;夏季处理A、处理C的芽头密度低于CK,与对照差异不显著(p>0.05),处理B、处理D、处理E的芽头密度高于CK,与对照差异显著(p<0.05),其中,处理B芽头密度最大;秋季处理C的芽头密度低于CK,其余处理均高于CK,秋季芽头密度无显著性差异(p>0.05)。综上可知,不同土壤改良措施对芽头密度显著性影响,但主要集中在春季和夏季,秋季无明显差异。表2 2022年不同季节芽头密度处理芽头密度/(个·尺–2) 春茶夏茶秋茶CK (46.33±3.06) b (46.67±1.53) bc (38.33±5.86) aA (51.00±2.65) a (44.33±2.52) bc (44.33±1.53) aB (51.33±1.53) a (56.33±1.53) a (40.67±6.66) aC (49.33±2.08) ab (42.67±2.52) c (36.67±4.73) aD (49.00±1.00) ab (48.33±3.51) b (40.33±5.51) aE (46.67±2.08) b (46.33±3.21) bc (38.33±5.69) a最小值 46.33 42.67 36.67 最大值51.33 56.33 40.67 均值(48.94±2.69) (47.44±4.98) (39.78±5.08)变异系数/% 5.50 10.50 12.77 注:1尺≈33.33 cm。2.3调酸措施对百芽干重的影响从表3可以看出,春季、夏季和秋季的百芽
2025年2月热带农业科学第45卷第2期20 干重变异系数依次为9.01%、6.27%、8.02%,秋季的百芽干重低于春季和夏季,春季处理A、E的百芽干重低于对照(CK),处理B、C、D高于CK,不同处理百芽干重与CK相比,差异不显著;夏季、秋季所有处理的百芽干重均高于CK。不同处理百芽干重均与CK差异不显著,认为调酸措施对百芽干重无显著性影响。表3 2022年不同季节百芽干重处理百芽干重/g 春茶夏茶秋茶CK (2.33±0.23) a (2.45±0.08) a (1.80±0.08) aA (2.28±0.03) a (2.58±0.11) a (1.91±0.12) aB (2.35±0.26) a (2.61±0.27) a (1.85±0.17) aC (2.38±0.20) a (2.58±0.11) a (1.91±0.13) aD (2.45±0.28) a (2.54±0.15) a (1.83±0.05) aE (2.18±0.24) a (2.51±0.25) a (1.95±0.31) a最小值 2.18 2.45 1.80 最大值2.45 2.61 1.95 均值(2.33±0.21) (2.55±0.16) (1.87±0.15)变异系数/% 9.01 6.27 8.02 2.4调酸措施对茶叶产量的影响比较2022年不同季节不同处理下的茶叶产量,结果见表4。由表4可知,所有处理的夏秋茶的鲜叶产量均高于春茶,春茶鲜叶产量变异系数为9.86%,处理D、处理E产量低于对照(CK),处理A、B、C鲜叶产量高于CK。其中,处理B的产量最高,为21.80 kg/亩(1亩≈667 m2),夏茶的变异系数为12.90%,所有处理鲜叶产量均高于CK,产量最高的是处理E,为46.83 kg/亩,秋茶的变异系数为14.93%,处理C的鲜叶产量低于表4 2022年不同季节鲜叶产量处理鲜叶产量/(kg·667 m–2) 春茶夏茶秋茶CK (18.90±2.51) a (42.17±5.14) a (41.70±8.46) aA (19.80±1.39) a (43.40±5.57) a (47.37±6.54) aB (21.80±1.85) a (45.47±1.44) a (46.07±4.65) aC (19.63±2.60) a (42.13±8.70) a (40.67±6.41) aD (18.83±1.20) a (42.77±9.38) a (42.00±8.19) aE (18.43±0.86) a (46.83±4.47) a (46.70±7.66) a最小值18.43 42.17 40.67 最大值 21.80 46.83 47.37 均值 (19.57±1.93) (43.79±5.65) (44.08±6.58)变异系数/% 9.86 12.90 14.93 CK,其余处理的鲜叶产量高于CK,其中,处理B的产量最高,为47.37 kg/亩,不同季节不同处理间差异不显著,说明在进行土壤酸化改良期间,对茶园的鲜叶产量产生影响较小。进行调酸措施后的年产量如图2所示。与对照(CK)相比,处理1年后处理A、E增产率分别为1.48%、9.27%,其余处理产量均有所下降;处理2年后处理A、B、E增产率分别为4.43%、4.22%、11.44%,处理C、D产量有所下降,处理E的产量较CK有明显增加,表明该处理在降低土壤酸性的同时,茶叶的产量也有所提高;2022年茶叶产量与2021年相比,除处理D外,各处理的茶叶产量均有增加,处理A、B、C、E的增产率分别为1.55%、8.20%、3.23%、0.60%,各处理产量无显著性差异。结合各处理的土壤pH可知,施用双飞粉能够在短期内显著提高茶园pH,但在2022年停止施用双飞粉后,处理B、C在2022年的pH显著降低,同时茶叶产量也有所提高。图2不同土壤改良措施的茶叶产量变化3讨论与结论3.1讨论适合茶树栽培的pH为4.0~6.5,最适合茶树栽培的pH为4.5~5.5[21]。通过在土壤中添加土壤改良剂能够提高土壤pH,修复酸化土壤[22-23]。2年酸化土壤改良期间,所有处理的pH均调整至适合茶树栽培的范围内,其中处理C、D、E的pH最适合茶树栽培。当处理中含有双飞粉时,能够在短时间内显著提高茶园土壤pH,这可能与土壤改良剂自身含有碱性物质有关,碱性物质能够中和土壤的酸根离子,进而提高土壤的pH[24]。处
刘春艳等不同调酸措施对酸化茶园土壤改良以及茶叶产量的影响-21 -理B、C在第2年取消双飞粉后,pH显著降低,而处理A依旧存在上升趋势,这可能是处理中微生物菌肥和有机肥的作用。吕秀娟等[25]研究表明,炭基微生物有机肥能提高一芽一叶的百芽重,加快茶叶的生长速度,显著提升茶叶品质。微生物菌肥可以改善土壤容重及透气性,提高养分利用效率[26]。与第1年相比,处理B增产显著,且第2年不同季节的茶叶产量、芽头密度、百芽干重均占优势,这可能与第二年施加了微生物菌肥有关。3.2结论对酸化茶园土壤采用不同的调酸措施,均能不同程度地提高茶园土壤的pH,处理期间在提高茶园pH的同时,茶叶年产量与CK差异不显著,说明对茶园进行调酸处理时,对茶叶产量产生影响较小,其中处理E的茶叶产量最高,说明该处理能够在酸化改良的过程中有一定提高茶叶产量的效果,处理B在第二年减施双飞粉后,pH下降显著,茶叶产量提高。不同处理芽头密度具有显著性差异,百芽干重不具有显著性差异,鲜叶产量春季和秋季处理B占优,夏季处理E占优;芽头密度春季和夏季处理B占优,秋季处理C占优;百芽干重春季处理D占优,夏季处理B占优,秋季处理E占优。综上可知,处理两年后,处理C、处理D、处理E均在最适合茶树栽培的pH范围内,其中,处理B在停施双飞粉后,pH下降,产量表现出一定的优势,处理E经过处理后,pH适中,且产量均占优势。参考文献[1]Lei W, Clayton R B, Qiuhui C, et al. Surface amendments can ameliorate subsoil acidity in tea garden soils of highrainfall environments[J]. Pedosphere,, 2016, 26(2). [2]张翔宇,尹彤云,周少奇,等. 中国植茶省份茶园土壤酸化现状分析[J]. 贵州科学, 2022, 40(3): 81-85. [3]Shiyu L, Haixia L, Changliang Y, et al. Rates of soil acidification in tea plantations and possible causes[J]. Agriculture, Ecosystems and Environment, 2016, 233. [4]Yan P, Wu L, Wang D, et al. Soil acidification in Chinese tea plantations[J]. Sci Total Environ, 2020, 715: 136 963. [5]樊战辉,唐小军,郑丹,等. 茶园土壤酸化成因及改良措施研究和展望[J]. 茶叶科学, 2020, 40(1): 15-25. [6]Jian Z, Ruidong Y, Yuncong C L, et al. Distribution, accumulation, and potential risks of heavy metals in soil and tea leaves from geologically different plantations[J]. Ecotoxicology and Environmental Safety, 2020, 195(C). [7]余跑兰,孙永明,李小飞,等施用白云石粉对茶园土壤酸化及茶叶品质与产量的影响[J]. 茶叶通讯, 2020, 47(1): 46-51. [8]L W, C R B, Y W, et al. Effect of crop residue biochar on soil acidity amelioration in strongly acidic tea garden soils[J]. Soil Use and Management, 2014, 30(1). [9]Zejiang C, Boren W, Minggang X, et al. Intensified soil acidification from chemical N fertilization and prevention by manure in an 18-year field experiment in the red soil of southern China[J]. Journal of Soils and Sediments, 2015, 15(2). [10]吴林土,徐火忠,李贵松,等. 微生物菌剂对松阳茶园土壤酸化改良及肥力提升的效果分析[J]. 浙江农业科学. 2022, 63(6): 1 245-1 249. [11]李艳春,汪航,李兆伟,等. 几种改良措施对酸化茶园土壤理化性质和微生物群落结构的影响[J]. 茶叶科学, 2022, 42(5): 661-671. [12]徐仁扣. 土壤酸化及其调控研究进展[J]. 土壤, 2015, 47(2): 238-244. [13]杨叶华,刘彦伶,李渝,等. 我国茶园土壤酸化阻控技术研究进展[J]. 现代园艺. 2023, 46(07): 39-40. [14]郭龙,李陈,刘佩诗,等. 牛粪有机肥替代化肥对茶叶产量、品质及茶园土壤肥力的影响[J]. 水土保持学报, 2021, 35(6): 264-269. [15]刘佩诗,黄瑜,甘曼琴,等. 茶园土壤有机肥施用效应和施肥技术[J]. 中国土壤与肥料, 2021(2): 306-311. [16]孟红旗,吕家珑,徐明岗,等. 有机肥的碱度及其减缓土壤酸化的机制[J]. 植物营养与肥料学报, 2012, 18(5): 1 153-1 160. [17]侯翠红,苗俊艳,谷守玉,等. 以钙镁磷肥产品创新促进产业发展[J]. 植物营养与肥料学报, 2019, 25(12): 2 162- 2 169. [18]余小兰,杨福锁,周丹,等. 钙镁磷肥对水稻土和砖红壤土壤化学性状的动态影响[J]. 江苏农业学报, 2018, 34(5): 1 042-1 047. [19]梁芳,涂卫佳,薛清华,等. 石灰和钙镁磷肥施用对水稻生长与镉累积的影响[J]. 湖南有色金属, 2018, 34(2): 56-60. [20]何燕燕. 微生物菌肥在农业生产中的价值和应用[J]. 工业微生物. 2023, 53(2): 8-10. [21]赖飞,张宝林,杨清,等凤冈县茶园土壤pH值变异特征及调控对策[J]. 农技服务, 2012, 29(5): 570-571. [22]Huang T H, Lai Y J, Hseu Z Y. Efficacy of cheap amendments for stabilizing trace elements in contaminated paddy fields[J]. Chemosphere, 2018, 198: 130-138.
2025年2月热带农业科学第45卷第2期22 [23]Ying H, Xu X, Xien L. Fungal denitrification contributes significantly to N2O production in a highly acidic tea soil[J]. Journal of Soils and Sediments, 2017, 17(6). [24]谢少华,宗良纲,褚慧,等. 不同类型生物质材料对酸化茶园土壤的改良效果[J]. 茶叶科学, 2013, 33(3): 279- 288. [25]吕秀娟,覃中凤,何志强,等. 炭基微生物有机肥施用量对覃塘毛尖茶百芽重及品质的影响[J]. 云南农业科技, 2023(3): 6-7. [26]决超. 微生物菌肥与土壤改良基质对连作马铃薯土壤性质及微生物群落的影响[J]. 江苏农业科学, 2023, 51(1): 218-224. (责任编辑龙娅丽)
2025年2月热带农业科学第45卷第2期Feb. 2025 CHINESE JOURNAL OF TROPICAL AGRICULTURE Vol.45, No.2 收稿日期 2024-03-16;修回日期 2024-05-08 基金项目 中央引导地方科技发展专项:海南白沙特色水果与茶叶绿色精准高效种植技术创新示范与推广应用(No. ZY2020 HN12);国家重点研发计划:“科技助力经济2020”重点专项——白沙地区热带果茶绿色精准高效种植技术应用与示范(No. SQ2020YFF0404782)。第一作者韩芳滨(1996—),女,硕士研究生,研究方向为农业昆虫与害虫防治,E-mail:kooros@163.com。通信作者 于广威(1975—),男,博士,教授,研究方向为病虫害绿色防控,E-mail:yuguangwei@139.com。海南白沙新建茶园主要害虫天敌种群与害虫消长动态分析韩芳滨1,2于广威3,4李超1(1. 新疆农业大学农学院/农林有害生物监测与安全防控重点实验室新疆乌鲁木齐 830052;2. 青岛罗素生物技术有限公司山东青岛 266104;3. 潍坊科技学院山东寿光 262700;4. 海南阿罗多甘农业科技有限公司海南白沙 572800)摘要为了更系统地探究海南白沙地区茶园中茶树的主要害虫和天敌的种类,以及害虫的发生规律,2023年全年对海南白沙一新建茶园内的害虫和天敌进行了系统的调查和研究。结果表明:茶园中有害的节肢动物群落物种主要由直翅目、蜚蠊目、半翅目、缨翅目、鞘翅目、双翅目、鳞翅目、蜱螨目组成;从种群数量来看,以半翅目为主,占害虫总数的43.66%;天敌以蜘蛛为主,占天敌总数的33.33%。共鉴定出茶树害虫为昆虫纲7目,42科69种,蛛形纲1目2科2种;鉴定出天敌:昆虫纲6目16科22种;蛛形纲1目,4科11种。在海南白沙的自然条件下4种害虫[小贯小绿叶蝉Empoasca (Matsumurasca) onukii Matsuda、茶蚜Toxoptera aurantii Boyer de Fonscolomb、茶黄蓟马Scirtothrips dorsalis Hood、绿盲蝽Apolygus lucorum Meyer-Dür]全年均发生,茶黄蓟马和绿盲蝽在上半年为害较严重,而小贯小绿叶蝉、茶蚜在下半年为害较严重。调查结果可为海南白沙的茶树虫害防治提供参考依据。关键词白沙;海南;茶树害虫;种群动态;调查中图分类号S533 文献标识码 A DOI: 10.12008/j.issn.1009-2196.2025.02.005 Analysis of the Population Dynamics of Major Pests and Their Natural Enemies in Newly Established Tea Plantations in Baisha, Hainan HAN Fangbin1,2 YU Guangwei3,4 LI Chao1(1. College of Agronomy, Xinjiang Agricultural University/Key Laboratory of the Pest Monitoring and Safety Control of Crops andForests, Urumqi, Xinjiang 830052, China; 2. Qingdao Russell Biotechnology Co., Ltd., Qingdao, Shandong 266104, China; 3. Weifang University of Science & Technology, Shouguang, Shandong 262700, China; 4. Hainan Alodogan Agricultural Science and Technology Limited Company, Baisha, Hainan 572800, China) Abstract A year-long investigation was carried out in 2023 in a newly established tea garden in Baisha, Hainan, to study the main pests and natural enemies of tea trees and the occurrence patterns of the pests. The results revealed that the arthropod community in the tea garden mainly comprises Orthoptera, Blattodea, Hemiptera, Thysanoptera, Coleoptera, Diptera, Lepidoptera, and Acari. Hemiptera was the predominant group, constituting 43.66% of the total pests, whereas spiders were the primary natural enemies, accounting for 33.33% of the total number of natural enemies. Seven insect orders with 42 families and 69 species, one arachnid order with two families and two species were identified as tea pests, Six insect orders with 16 families and 22 species, and one arachnid order with four families and 11 species as natural enemies. In Baisha, Hainan, four pest species, namely Empoasca (Matsumurasca) onukii Matsuda, Toxoptera aurantii Boyer de Fonscolomb, Scirtothrips dorsalis Hood, and Apolygus lucorum Meyer-Dür, are present year-round. Scirtothrips dorsalis Hood and Apolygus lucorumMeyer-Dür cause significant damage in the first half of the year. However, Empoasca (Matsumurasca) onukii Matsuda and Toxoptera aurantii Boyer de Fonscolomb cause more significant damage in the second half. These findings will contribute to pest control references for tea trees in Baisha, Hainan. Keywords Baisha; Hainan; tea tree pests; population dynamics; investigation
2025年2月热带农业科学第45卷第2期24 我国是茶叶的原产地,海南作为中国茶叶的主要产区之一,是全国唯一可以四季采茶的地区。海南茶区全年平均气温在23℃以上,一年内最冷月平均气温也在15℃以上。降水量丰富,年降水量为1 600~2 000 mm,适合茶树生长,因地理位置的特殊性,海南茶产业具有极大的发展潜力。得天独厚的地理自然条件也导致了害虫种类多,在此地区种植的茶树相比其他有明显昆虫越冬期地区的茶树更易受到害虫侵袭。对于海南白沙茶树害虫种群的研究,除多年前报道过的海南省茶树害虫名录[1-2],其中对海南白沙的茶树害虫种群有部分描述以外,未见更为详细的白沙地区的茶树害虫天敌种类调查和主要害虫的种群消长动态报告。为补充此部分数据,本研究对海南白沙的新建茶园进行为期一年茶树害虫天敌种群的调查,同时总结4种茶树主要害虫田间种群数量动态及规律,为海南白沙地区茶园的绿色防控提供参考依据。1材料与方法1.1材料试验地块设在海南省白沙黎族自治县阿罗多甘共享农庄的品种茶园内,气候温润、雨量丰沛,其茶园生态环境在海南茶区具有代表性。实验茶树品种为中茗7号和福云595,于2020年扦插,2022年1月定植,至2023年12月实验结束树龄为3年,均处于幼年期,福云595树高约57 cm,蓬面宽约40 cm,株距约35 cm,行距约150 cm;中茗7号树高约30 cm,蓬面宽约25 cm,株距约30 cm,行距约150 cm。1.2方法2023年1月至2023年12月在试验地块用五点取样法,每个小区中每点取边长取1 m的茶行6~8株茶树,每个品种各10个样点,共计20个样点。1.2.1目测调查法对于常栖息在叶片上的茶树害虫采用检叶数虫法,随机选取1 m茶行中6株茶树调查百叶,调查叶片为芽下二叶或三叶;对于常为害枝条和根部的害虫选取该范围内的茶树上、中、下层的样枝进行普查,所查样枝长度范围为10 cm;对所查茶树根据被害部位分类记录。每7 d调查1次,调查时间为清晨露水未干时,偶遇暴雨、降雪等不可抗拒因素,适当提前或者推迟调查时间。1.2.2 黄板观察法在样点附近设置20 cm× 25 cm的黄色粘板,每7 d更换1次黄板。将已用黄板用保鲜膜封住带入实验室计数和辨识昆虫。1.2.3室内观察法在每个调查点选取一株茶树,按上、中、下3层取样,每层取6个叶片放进自封袋中,并对其做好序号标记,带回试验室用体视显微镜进行观察和鉴定。1.2.4分类与鉴定在系统调查过程中,直接记录已知的害虫及其天敌,同时,对于未知的昆虫,需进行统一标注并带回实验室,然后通过体视显微镜观察其特征来进行识别。根据其外部特征不能鉴定出种类的幼虫,饲养至成虫后再进行鉴定。对常见且特征明显的个体鉴定到种,其他鉴定到科。以下为昆虫鉴定的参考文献及书籍:《昆虫分类学》《普通昆虫学》《茶树病虫及天敌图谱》《茶树病虫和天敌名录》等。1.2.5数据分析利用Excel 2017数据进行数据整合和作图。2结果与分析2.1茶园害虫及天敌的数量特征白沙新建茶园中有害的节肢动物群落物种主要由直翅目、蜚蠊目、半翅目、缨翅目、鞘翅目、双翅目、鳞翅目、蜱螨目组成。从种群的数量来看,以半翅目最多,占总数的43.66%,叶蝉在半翅目中种类最多;其次是缨翅目和鞘翅目,均占总种数的16.90%,直翅目占比为15.49%;蜚蠊目、双翅目、蜱螨目种类数占比最少(表1、图1)。茶园中的天敌主要由半翅目、脉翅目、膜翅目、鞘翅目双翅目、螳螂目、蜘蛛目所组成。其中,蜘蛛目种类数最为丰富,占天敌总数的33.33%,蜘蛛目天敌的食性杂,分为游猎型和定居型,可捕食小型害虫也可捕食大型害虫;其次为鞘翅目,占总数的18.18%,鞘翅目中的天敌主要为瓢虫科,它们专捕食小型的刺吸式害虫,例如茶蚜、蓟马、粉虱、叶蝉等,茶蚜在茶园中泛滥时瓢虫的数量也肉眼可见的增长。膜翅目与双翅目占比大致相同,主要为寄生蜂、捕食性蜂和食蚜蝇。茶园中的半翅目天敌主要为捕食性蝽,占比为12.12%,主要以刺吸其同类害蝽和
韩芳滨等海南白沙新建茶园主要害虫天敌种群与害虫消长动态分析-25 -其他害虫的体液为生。螳螂目与脉翅目天敌在茶园中种类较少,分别为天敌总种数的3.03%(表1、图2)。表1茶园害虫种类数量特征害虫(螨) 科种类数天敌科种类数直翅目Orthoptera 8 11 半翅目Hemiptera 4 4 蜚蠊目Blattaria 3 3 脉翅目Neuroptera 1 1 半翅目Hemiptera 14 31 膜翅目Hymenoptera 4 5 缨翅目Thysanoptera 2 3 鞘翅目Coleoptera 3 6 鞘翅目Coleoptera 8 12 双翅目Diptera 3 5 双翅目Diptera 2 2 螳螂目Mantodea 1 1 鳞翅目Lepidoptera 5 7 蜘蛛目Araneae 4 11 蜱螨目Arachnoidea 2 2 合计20 33 合计44 71 2.2茶园害虫及天敌的种类2023年1—12月在茶园害虫共鉴定出昆虫纲7目,42科,69种;鉴定出蛛形纲1目,2科,2种。鉴定出的茶园害(螨)虫及天敌见表2、3。图1茶园主要害虫种类占比图2茶园主要天敌种类占比表2海南白沙新建茶园主要害虫种类纲目科种名及拉丁名危害部位昆虫纲Insecta 蜚蠊目Blattodea白蚁科 Termitidae黑翅土白蚁Odontotermes formosanus Shiraki 根部鼻白蚁科Rhinotermitidae 台湾家白蚁Coptotermes formosanus Shiraki 根部姬蠊科Ectobiidae 姬蠊Blattella sp.啃咬叶片直翅目Orthoptera 蝗科Acrididae 中华剑角蝗Acrida cinerea Thunb 食叶中华稻蝗Oxya chinensis Thunb 食叶斑翅蝗科Oedipodidae 疣蝗Trilophidia annulata Thunberg食叶锥头蝗科Pyrgomorphidae 短额负蝗Atractomorpha sinensis Bolivar 食叶斑腿蝗科Catantopidae 外斑腿蝗Xenocatantops Dirsh 食叶蟋蟀科Grylloidea 油葫芦Cryllus testaceus Wallker 根、茎、叶树蟋科Oecanthinae 中华树蟋Oecanthus indicus Saussure 根、茎、叶蛉蟋科Trigonidiidae 虎甲蛉蟋Trigonidium cicindeloides Rambur 根、茎、叶斑腿双针蟋Dianemobius fascipes Walker 根、茎、叶螽斯科Tettigonioidea 绿螽斯Holochlora nawae Mats.et Shiraki嫩芽、嫩叶悦鸣草螽Conocephalus melaenus Haan 嫩芽、嫩叶
2025年2月热带农业科学第45卷第2期26 续表2海南白沙新建茶园主要害虫种类纲目科种名及拉丁名危害部位昆虫纲Insecta 半翅目Hemiptera 蚜科Aphidoidea 茶蚜Toxoptera aurantii Boyer de Fonscolomb 嫩芽、嫩叶蝽科Pentatomoidea 珀蝽Plautia fimbriata Fabricius 刺吸叶片稻赤曼蝽Menida versicolor Gmelin 刺吸叶片盲蝽科Miridae 绿后丽盲蝽(绿盲蝽)Apolygus lucorum Meyer-Dür 网蝽科Tingidae 茶脊冠网蝽(茶网蝽)Stephanitis chinensis Drake 刺吸叶片缘蝽科Coreidae 稻棘缘蝽Cletus punctiger Dallas 刺吸叶片中稻缘蝽Leptocorisa chinensis Dallsa 刺吸叶片条蜂缘蝽Riptortus linearis Fabr 刺吸叶片长蝽科Lygaeidae 红脊长蝽Tropidothorax elegans Distant 刺吸叶片小长蝽Nysius ericae Schilling 刺吸叶片紫黑刺胫长蝽Horridipamera nietneri Dohrn 刺吸叶片地长蝽科Rhyparochromidae白斑地长蝽Rhyparochromus albomaculatus J. Scott 刺吸叶片龟蝽科Plataspidae 龟蝽Plataspidae刺吸叶片叶蝉科Cicadellidae 小贯小绿叶蝉Empoasca (Matsumurasca) onukii Matsuda 嫩叶、芽梢电光叶蝉Recilia dorsalis Motschulsky 嫩叶、芽梢二条黑尾叶蝉Nephotettix nigropictus Motschulsky 嫩叶、芽梢黑颜单突叶蝉Lianodiana brevis Cai & He 嫩叶、芽梢白翅褐脈大叶蝉Cofana spectra Distant 嫩叶、芽梢茶扁叶蝉Penthimiatestacea Kuoh 嫩叶、芽梢片头叶蝉Petalocephala Stal 嫩叶、芽梢菱纹叶蝉Hishimonus sp.嫩叶、芽梢顶带叶蝉Exitianus sp.嫩叶、芽梢角蝉科Membracidae 白翅三刺角蝉Tricentrus albipennis Kato 嫩叶、芽梢短屈角蝉Anchon brevicornis Chou嫩叶、芽梢茶树三刺角蝉Tricentrus camelliae Yuan et Fan 嫩叶、芽梢黑圆角蝉Gargara genistae Fabricius 嫩叶、芽梢蜡蝉科Fulgoridae 八点广翅蜡蝉Ricania speculum Walker 芽、叶象蜡蝉科Dictyopharidae 中华彩象蜡蝉Raivuna sinica Walker 芽、叶粉虱科Aleyrodidae 白粉虱Trialeurodes vaporariorum Westwood叶黑刺粉虱Aleurocanthus camelliae Kanmiya&Kasai 叶粉蚧科Pseudococcidae 粉蚧Pseudococcus 叶、枝干缨翅目Thysanoptera 蓟马科Thripidae 茶黄蓟马Scirtothrips dorsalis Hood 嫩叶茶棍蓟马Dendrothrips minowai Priesner 嫩叶管蓟马科Phlaeothripidae 榕管蓟马Gynaikothrips uzeli Zimmerman 嫩叶鞘翅目Coleoptera 天牛科Cerambycidae 茶天牛Aeolesthes induta Newman 幼虫蛀食枝干、根部叶甲科Chrysomelidae 黄斑长附萤叶甲Monolepta signata Olivier 嫩梢、芽叶褐背小萤叶甲Galerucella grisescens Joannis 嫩梢、芽叶黑足田守瓜Aulacophora nigripennis Motschulsky 嫩梢、芽叶黄曲条跳甲Phyllotretastriolata Fabricius 嫩梢、芽叶金龟科Scarabaeidae 中喙丽金龟Adoretus sinicus Burmeister 根部、叶片黄金龟甲Charidotella sexpunctata Fabricius 根部、叶片象甲科Curculionidae 茶芽粗腿象甲Ochryromera quadrimaculata Voss 嫩叶
韩芳滨等海南白沙新建茶园主要害虫天敌种群与害虫消长动态分析-27 -续表2海南白沙新建茶园主要害虫种类纲目科种名及拉丁名危害部位昆虫纲Insecta 鞘翅目Coleoptera 叩甲科Elateridae 叩甲Elateridae幼虫蛀食枝干、根部拟步甲科Tenebrionidae 拟步甲Tenebrionidae芽、茎皮蠹科Dermestidae 圆皮蠹Anthrenus Schaeffer 种子小蠹科Scolytinae 茶枝小蠹Euwallacea fornicatus Eichhoff 蛀食枝干双翅目Diptera 实蝇科Tephritidae 橘小实蝇Bactrocera dorsalis Hendel 果实、种子潜蝇科Agromyzidae 茶潜叶蝇Chlorops theae Lefroy 幼虫食叶肉鳞翅目Lepidoptera 刺蛾科Limacodidae 茶刺蛾Iragoides fasciata Moore食叶鹿蛾科Ctenuchidae 蕾鹿蛾Amata germana Felder 食叶卷蛾科Tortricidae 茶小卷叶蛾Adoxophyes orana Fischer von Rosl 食叶尺蛾科Geometridae 茶尺蠖Ectropis oblique hypulina Wehrli 食叶油桐尺蠖Buasra suppressaria Guenee 食叶蓑蛾科Psychidae 茶蓑蛾Clania minuscula Butler食叶茶小蓑蛾Acanthop syche sp.食叶蛛形纲Arachnida 蜱螨目Acarina 跗线螨科Tarsonemidae 茶黄螨Polyphagotarsonemus latus Banks 刺吸叶片细须螨科Tenuipalpidae 茶短须螨Brevipalpus obovatus Donnadieu 刺吸叶片表3海南白沙新建茶园主要天敌种类纲目科种名及拉丁名主要取食/寄生害虫昆虫纲Insecta 鞘翅目Coleoptera 瓢虫科Coccinellidae 六斑月瓢虫Cheilomenes sexmaculata Fabricius 蚜虫、蓟马、叶蝉、龟纹瓢虫Propylea japonica Thunberg 粉虱等异色瓢虫Harmonia axyridis Pallas 蚜虫、蓟马、叶蝉等红点唇瓢虫Chilocorus kuwanae Silvestri 蚧壳虫步甲科Carabidae 步甲Carabidae鳞翅目幼虫、蚜虫等虎甲科Cicindelidae 虎甲Cicindelidae食性广泛,螨、鳞翅目幼虫、蟋蟀等半翅目Hemiptera 花蝽科Anthocoridae 小花蝽Orius similis Zheng 蚜虫、蓟马、螨等猎蝽科Reduviidae 红彩瑞猎蝽Rhynocoris fuscipes Fabricius 鳞翅目幼虫等长蝽科Lygaeidae 宽大眼长蝽Geocoris varius Uhler 蚜虫、蓟马、盲蝽螳螂目Mantodea 花萤科Cantharidae 短翅花萤Trypherus sp. 蚜虫、蛾类幼虫螳科Mantidae 中华大刀螳Tenodera sinensis Hermann Burmeister 食性广泛脉翅目Neuroptera 草蛉科Chrysopidae 中华大草蛉Chrysopa pallens Rambur 蚜虫、螨等双翅目Diptera 食蚜蝇科Syrphidae 钝黑斑眼蚜蝇Eristalinus sepulchralis Linnaeus 蚜虫亮黑斑眼蚜蝇Eristalinus tarsalis Macquart 蚜虫黄跗斑眼蚜蝇Eristalinus quinquestriatus Fabricius 蚜虫麻蝇科Sarcophagidae 麻蝇Sarcophagidae鳞翅目幼虫寄蝇科Tachinidae 寄蝇Tachinidae鳞翅目幼虫、蝽膜翅目Hymenoptera 茧蜂科Braconidae 茧蜂Braconidae鳞翅目幼虫姬蜂科Ichneumonidae 姬蜂Ichneumonidae鳞翅目幼虫小蜂科Chalcidoidea 小蜂Chalcidoidea鳞翅目幼虫
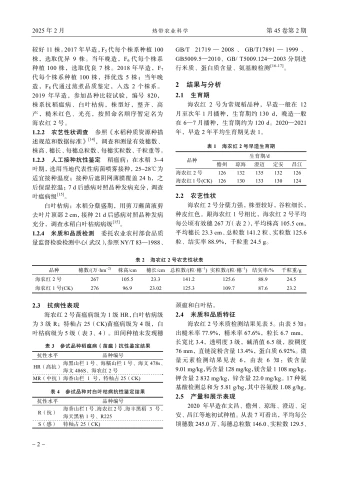
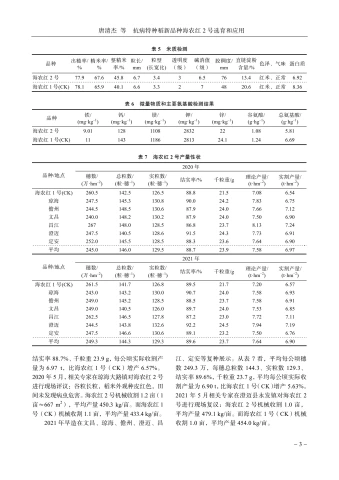
2025年2月热带农业科学第45卷第2期28 续表3海南白沙新建茶园主要天敌种类纲目科种名及拉丁名主要取食/寄生害虫昆虫纲Insecta 膜翅目Hymenoptera 胡蜂科Vespidae 胡蜂Vespidae鳞翅目、半翅目、鞘翅目等体型较大的害虫蜾蠃Eumeninae鳞翅目幼虫蛛形纲蜘蛛目Araneae 跳蛛科Salticidae 条纹金蝉蛛Phintella vittata Karsch 叶蝉、蚜虫、飞虱、蚊蝇等小型害虫白斑艾普蛛Epeus alboguttatus Thorell双尖艾普蛛Epeus bicuspidatus Song,Guet Chen多彩纽蛛Telamonia festiva Thorell 角猫跳蛛Carrhotus sannio Thorell 阔莎茵蛛Thyene imperialis Rossi叶蝉、蚜虫、飞虱、蚊蟹蛛科Thomisidae 角红蟹蛛Thomisus labefactus Karsch 蝇等小型害虫园蛛科Araneidae 横纹金蛛Argiope bruennichi Scopoli 食性广泛,小到叶蝉、蚜虫、蚊蝇大到蟋蟀、蝗虫等猫蛛科Oxyopidae 台湾绿猫蛛Peucetia formosensis Kishida 细纹猫蛛Oxyopes macilentus L.Koch 斜纹猫蛛Oxyopes sertatus L.Koch 天敌共鉴定出昆虫纲6目,16科,22种;蛛形纲1目,4科11种。鉴定出的茶园天敌如下。2.3茶园主要害虫种群消长动态分析2023年112月对茶园中种植面积最大的茶树品种福云595和中茗7号,分别取10个样点进行调查,对发生量大的4种害虫——小贯小绿叶蝉Empoasca (Matsumurasca) onukii Matsuda、茶蚜Toxoptera aurantii Boyer de Fonscolomb、茶黄蓟马Scirtothrips dorsalis Hood、绿盲蝽Apolygus lucorum Meyer-Dür进行种群消长动态分析。2.3.1小贯小绿叶蝉种群数量动态小贯小绿叶蝉喜呆叶背阴凉处,个体小世代重叠严重,主要通过刺吸茶树嫩芽、叶的汁液来对茶树造成伤害,受害部位萎缩,叶缘和叶尖会呈现红褐色枯萎为害症状,导致生长停止[3]。该虫于2月下旬开始出现,大部分形态为若虫。2个品种的虫害发生规律基本相似,共出现4个高峰:分别为3月中旬、4月上旬、5月中旬和10月下旬(图3)。其中,2月下旬至3月中旬,小贯小绿叶蝉虫口数量开始增加,福云595达到120头,中茗7号最高达到85头,此时茶园红褐色叶片明显变多;3月下旬至6月期间虽有小波动但整体呈现下降的趋势;9月至11月的海南白沙阴雨天为主,此时小贯小绿叶蝉发生量明显多于前期。从9月虫口数开始上升,直到10月下旬达到最高峰,福云595达到448头,中茗7号最高达到551头。后茶园进行定期修剪工作,虫口数呈下降趋势,11月下旬虫口数开始回升,此时福云595达到了166头,中茗7号最高达160头,达到另一个峰值,但较前一次高峰虫口数有所减少。2.3.2茶蚜种群数量动态在茶蚜为害高峰期时可以观察到茶蚜成、若虫聚集在茶叶新梢嫩叶或茎的背面,为害较严重时也可以观察到其聚集在芽头,使得芽叶细弱、卷缩并排泄“蜜露”使茶叶染病。从1月调查开始茶蚜虫口数量就达到了第一个高峰,其中福云595为1 122头,中茗7号889头。4月上旬,从图4可看出,福云595达到第二个高峰363头,而中茗7号稍微晚一些,于4月中旬达到第二个高峰428头,8月上旬达到第三个峰值381头,此时福云595受茶蚜为害小,但8月下旬开始,茶蚜虫口数量开始急剧上升,于9月中下旬达到全年茶蚜最大发生量3 593头。而此时中茗7号达到了第4个高峰,但茶蚜发生量少于福云595为1 101头,10月上旬达到第5个高峰673头,而福云595此时茶蚜数开始上升直到10月下旬达到最后一个高峰1 701头,随后呈现缓慢下降趋势,而中茗7号呈现上升趋势。至调查的最后时间12月中旬,福云595为578头,中茗7号865头。两品种茶蚜发生数量和趋势出偶见差别,大部分时间基本相似,数量相差不大。
韩芳滨等海南白沙新建茶园主要害虫天敌种群与害虫消长动态分析-29 -图3小贯小绿叶蝉种群数量消长动态图4茶蚜种群数量消长动态2.3.3茶黄蓟马种群数量动态茶黄蓟马的成、若虫主要为害茶树的嫩梢和嫩叶。从图5可看出,两品种的茶黄蓟马种群数量消长动态全年基本一致,虫口数从1月初开始上升,春季3月上旬达到了全年最高峰,福云595为4 331头,中茗7号为4 377头,春茶受害严重。3月中旬茶黄蓟马的虫口数整体呈现下降的趋势,期间4月18日中茗7号又迎来一波小高峰,虫口数量为1 870头。6月下旬至7月初,福云595和中茗7号达到另一个峰值,分别为924和1 524头,此期间中茗7号
2025年2月热带农业科学第45卷第2期30 的峰值稍晚一些。6月下旬至9月上旬,中茗7号共达到3个峰值,分别为6月30日的1 524头,8月7日的1 269头和9月5日的1 280头,而福云595为2个高峰,分别为6月24日的924头和8月27日的1 392头。9月中旬开始,茶黄蓟马数量开始下降,9月下旬虽有回升但数量小于前2个时期。10月17日后茶黄蓟马的虫口数不再上升。2.3.4绿盲蝽种群数量动态绿盲蝽在田间通常昼伏夜出,在该茶园春季羽化后迁出,后少有发生。从图6可看出,福云595的绿盲蝽为害期主图5茶黄蓟马种群数量消长动态图6绿盲蝽种群数量消长动态
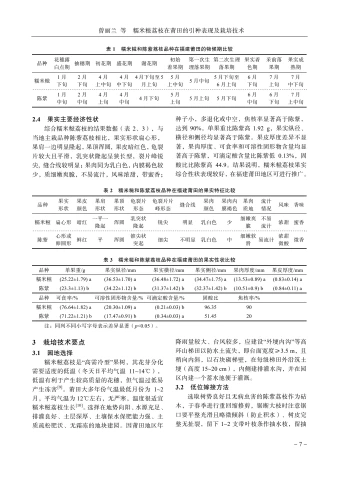
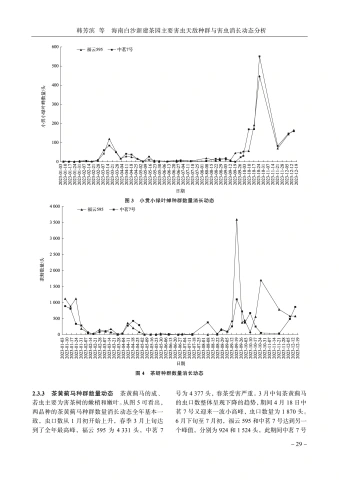
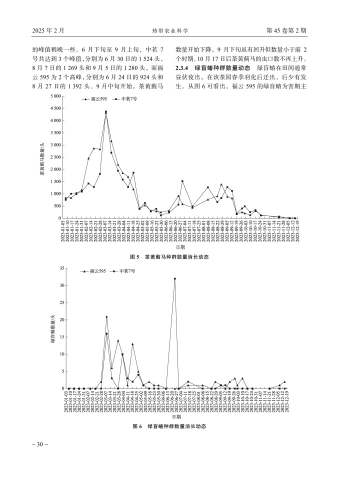
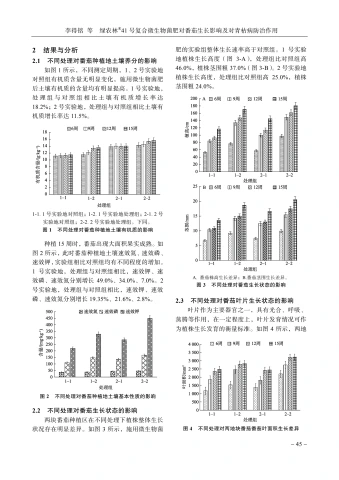
韩芳滨等海南白沙新建茶园主要害虫天敌种群与害虫消长动态分析-31 -要在2月下旬至4月下旬,这期间共出现了3个高峰,其中3月上旬达到了最大值,福云595为21头、中茗7号16头。6月中旬至下旬,中茗7号受绿盲蝽为害严重,达到全年最大值32头,而福云595此时受绿盲蝽为害最小。2个品种后续7月中旬、8月下旬至10月中旬绿盲蝽种群有小起伏,但种群数量都不超过3头。除6月中下旬期间,其他时间2种茶树绿盲蝽种群动态相似,被绿盲蝽为害的茶叶呈现“破叶疯”症状:当食用部位受到破坏时,通常会形成坏死点并留下一个孔洞。随着受害植物的生长,这些孔洞可能会逐渐扩大并呈现不规则形状,叶片可能会出现破损或残缺。受害区域的边缘则会逐渐褪绿、变黄,并且厚度增加[4],严重影响了春茶生产。3讨论与结论茶园中的害虫与天敌群落构成的生态系统对茶树生长具有重要的影响。从研究结果可以看出,白沙新建茶园中存在着丰富多样的节肢动物群落,其中害虫以半翅目为主,而半翅目害虫包括了叶蝉、茶蚜、绿盲蝽等主要刺吸式口器害虫,与我国茶园现今由鳞翅目食叶类害虫向小型刺吸式害虫演替相一致[5]。茶园内的天敌以蜘蛛、瓢虫、寄生蜂和食蚜蝇等为主,蜘蛛目作为茶园中的重要天敌其狩猎范围很大,不仅捕食小型害虫,如茶蚜、叶蝉等,还能够捕食大型害虫,如鳞翅目害虫、蝗虫等。瓢虫和食蚜蝇的幼虫主要以茶蚜为食,寄生蜂则主要针对鳞翅目害虫。通过调查可知,茶园中的茶树害虫(螨)共鉴定出昆虫纲7目,42科,69种;鉴定出蛛形纲1目,2科,2种;天敌共鉴定出昆虫纲6目,16科,22种;蛛形纲1目,4科11种(表2、表3)。根据已记录海南茶树害虫相关文献对比[1-2],分布在琼中(岭头)的茶小卷叶蛾Adoxophyes oranaFischer von Rosl、茶小蓑蛾Acanthopsyche sp.等在此次对白沙新建茶园的调查中也有发现,其他普遍发生的茶树主要害虫,如小贯小绿叶蝉、茶黄蓟马、茶蚜、茶潜叶蝇等均出现。通过对海南省白沙黎族自治县阿罗多甘共享农庄新建茶园进行主要茶树害虫的消长动态调查,初步明确了小贯小绿叶蝉Empoasca (Matsumurasca) onukii Matsuda、茶蚜Toxoptera aurantiiBoyer de Fonscolomb、茶黄蓟马Scirtothrips dorsalis Hood、绿盲蝽Apolygus lucorum Meyer-Dür,4种害虫在茶园的种群数量消长动态。整体而言,4种害虫在福云595与中茗7号上的全年发生趋势基本一致。海南的春茶生产时间比内陆早,且未遭受霜冻和昆虫的侵扰,因此,它的茶叶收获期比中国其他地方提前1~3个月。海南的明前茶因此被誉为“华夏第一早春茶”[6]。1月正是海南春茶上市之时,此时茶树嫩梢、嫩叶多,茶蚜发生危害严重。该茶园小贯小绿叶蝉种群发生高峰期分别为3月中旬至5月中旬和9下旬至11月下旬,11月下旬后虫口数量短暂减少,随后12月继续上升,全年最大值都集中在下半年1011月,这点与贵州黔中、河南信阳地区相似,全年发生持续时间与浙江金华、江西遂川、广西梧州等地一致[7-9]。茶黄蓟马全年都有发生,且为害最严重的时期为3月中旬,被害叶片背面主脉两侧出现2条或数条褐色凹陷的纵纹,纵纹相应部位的叶正面隆起。当叶片遭受损害时,则向背部弯曲或者边缘变得不规则、扭曲、硬化,而新生的枝条或者叶的柄部呈现出褐色的斑点[10]。11月下旬,海南白沙地区开始降温,最低温度为12℃,此时受到短时低温的影响,虫口数量逐渐减少。在22~30℃的条件下,茶黄蓟马的发育历期缩短,由于海南白沙等地区冬季平均气温极少出现10℃及以下持续低温天气,所以相对于有明显越冬期的地区,应全年监测茶黄蓟马的动态,并制定适宜的防治策略[11-13]。绿盲蝽主要危害时间为春季,海南春季光照充足,气温在23℃上下。在此条件下,绿盲蝽的成活率和种群增长指数均达到最高水平。而绿盲蝽的生长发育繁殖受光周期、温度和湿度影响很大,长光照更有利于卵的孵化。而在短光照的作用下绿盲蝽多次交尾,以此来增加产卵量,白沙地区的雨量充足,而多雨的天气也增加了绿盲蝽卵的孵化率。绿盲蝽的各个发育阶段的起点温度都在10℃以下,这表明它们能够在较低的温度下进行发育[14-18]。因此掌握绿盲蝽的发生动态,做好防控预警工作能够极大地防止春季茶树“破叶疯”的症状。随着全球化趋势愈演愈烈,我国茶叶出口的
2025年2月热带农业科学第45卷第2期32 优势面临前所未有的挑战,为应对这一挑战,充分发挥和利用海南白沙地区独特的气候和环境优势,可促进茶产业的发展。其中,了解茶园内害虫天敌的种群以及主要害虫的数量动态至关重要。通过有效地调查和监测,制定相应的绿色防控策略,提升白沙茶的品质和市场竞争力,同时为国内茶产业的可持续发展奠定坚实基础。参考文献[1]王纪文. 海南茶树害虫名录[J]. 海南大学学报(自然科学版), 1986(1): 33-41. [2]赵冬香, 高景林, 王爱萍, 等. 海南省茶树害虫名录[C]//中国茶叶学会, 台湾茶协会. 第六届海峡两岸茶业学术研讨会论文集(摘要). 中国热带农业科学院环境与植物保护研究所, 2010: 1. [3]张君岱, 涂修亮, 毛迎新,等. 中国茶树小贯小绿叶蝉研究进展[J]. 湖北农业科学, 2017, 56(12): 2204-2208+ 2 211. [4]郭华伟. 恼人的游击者——绿盲蝽[J]. 中国茶叶, 2019, 41(2): 7-9. [5]陈宗懋, 蔡晓明, 周利, 等. 中国茶园有害生物防控40年[J]. 中国茶叶, 2020, 42(1): 1-8. [6]于伟慧. 2019中国(海南)早春茶开采季仪式启动, 海南春茶迈开大步迎接市场: 尝一尝海南春天的味道[J]. 世界热带农业信息, 2019(1): 47-49. [7]乔利, 吕立哲, 吴淑平, 等. 温度对豫南地区茶园小绿叶蝉种群的影响[J]. 中国农学通报, 2016, 32(1): 146-149. [8]杨春, 孟泽洪, 李帅, 等. 贵州茶园茶棍蓟马和小贯小绿叶蝉种群动态及主栽茶树品种寄主抗性[J]. 南方农业学报, 2021, 52(3): 671-681. [9]曹馨月, 王迎春, 龚雪蛟, 等. 小贯小绿叶蝉区域性发生规律和综合防治[J]. 中国植保导刊, 2018, 38(12): 25-32. [10]林雄毅. 茶黄蓟马的发生特点与防治方法[J]. 中国植保导刊, 2007(11): 28-29. [11]韩冬银, 邢楚明, 李磊, 等. 短时低温对茶黄蓟马发育与繁殖的影响[J]. 环境昆虫学报, 2023, 45(3): 666-672. [12]韩冬银, 李磊, 张方平, 等. 茶黄蓟马低温致死中温度、中时间测定[J]. 热带农业科学, 2023, 43(12): 65-68. [13]黄丽莉, 何海敏, 阙海勇, 等. 茶黄蓟马的生物学特性[J]. 生物灾害科学, 2013, 36(3): 247-250. [14]卓德干, 李照会, 门兴元,等. 低温和光周期对绿盲蝽越冬卵滞育解除和发育历期的影响[J]. 昆虫学报, 2011, 54(2): 136-142. [15]赵洪霞, 肖留斌, 谭永安, 等. 不同光周期对绿盲蝽实验种群生命表参数的影响[J]. 棉花学报, 2011, 23(2): 140- 146. [16]PAN Hongsheng, LIU Bing, LU Yanhui, et al. Identification ofthe key weather factors affecting overwintering success of Apolygus lucorum eggs in dead host tree branches[J/OL]. PLoS ONE. 2014, 9(4): e94190. DOI: 10.1371/journal.pone. 0094190. [17]MEN Xing-yuan, Yu Yi, Zhang An-sheng, et al. Life table of the laboratory population Apolygus lucorum Meyer-Dür (Hemiptera: Miridae) at different temperatures[J]. Acta Entomologica Sinica, 2008, 51(11): 1 216-1 219. [18]朱秀红, 陈志超, 于怀征. 日照茶区主要虫害发生规律、适宜气象指标及防治技术[J]. 农业工程, 2022, 12(7): 135- 139. (责任编辑龙娅丽)
2025年2月热带农业科学第45卷第2期Feb. 2025 CHINESE JOURNAL OF TROPICAL AGRICULTURE Vol.45, No.2 收稿日期 2024-04-07;修回日期 2024-05-22 基金项目 海南省自然科学基金项目“硬枝黄蝉中环烯醚萜类化合物对辣椒疫霉菌的抑菌机理研究”(No.321MS092);海南省自然科学基金项目“诺丽提取物对咖啡炭疽病菌抑菌活性研究”(No.322QN401)。第一作者苟亚峰(1980—),女,硕士,副研究员,主要研究方向为植物源农药研究开发,E-mail:gouyf2007@163.com。通信作者 孙世伟(1981—),男,硕士,副研究员,主要研究方向为热带香料饮料作物病虫害防控,E-mail:sunshw@126.com。硬枝黄蝉中4种环烯醚萜类化合物对辣椒疫霉菌的抑菌活性研究苟亚峰1,2田甜1温思为1刘世超1高圣风1孙世伟1(1. 中国热带农业科学院香料饮料研究所海南万宁 571533;2. 海南省特色热带作物适宜性加工与品质控制重点实验室海南万宁 571533)摘要环烯醚萜类化合物作为众多植物中的有效成分之一,具有广泛的生物活性,如抗病毒、抗菌、抗肿瘤、抗氧化等,前期从硬枝黄蝉中分离得到4种环烯醚萜类化合物:plumieride、protoplumericin A、13-dexoylplumieride和13-O-p-coumaroylplumieride。采用生长速率法测定了上述4种化合物对辣椒疫霉菌的抑菌活性,并通过扫描电镜观察抑菌活性化合物对辣椒疫霉菌表观形态的影响。结果表明,供试化合物中protoplumericin A和13-O-p-coumaroylplumieride对辣椒疫霉菌具有一定抑菌活性,抑制率分别达到了72.97%和69.92%;采用13-O-P-coumaroylplumieride和protoplumieride A处理后,菌丝出现不规则收缩,表面塌陷或模糊等症状。研究为进一步研制防治辣椒疫霉菌的植物源杀菌剂提供支撑。关键词硬枝黄蝉;辣椒疫霉菌;环烯醚萜类;抑菌活性中图分类号S436.418.1 文献标识码 A DOI: 10.12008/j.issn.1009-2196.2025.02.006 Study on the Antifungal Activity of Four Iridoids from Allemanda neriifolia Against Phytophthora capsiciGOU Yafeng1,2 TIAN Tian1 WEN Siwei1 LIU Shichao1 GAO Shengfeng1 SUN Shiwei1(1. Research Institute of Spice-and-Beverage Crops, Chinese Academy of Tropical Agricultural Sciences, Wanning, Hainan 571533, China; 2. Key Laboratory of Processing-Suitability and Quality Control of the Special Tropical Crops of Hainan Province, Wanning, Hainan 571533, China) Abstract As components of many plants, iridoids have a wide range of biological activities, such as antiviral, antibacterial, anti-tumor, antioxidant, etc. Four iridoids of plumieride, protoplumericin A, 13-dexoylplumieride and 13-O-p-coumaroylplumieride have been isolated from Allemanda neriifolia. The antifungal activity against Phytophthora capsici was measured via the growth rate method, and the changes in the morphology of P. capsici were observed by scanning electron microscopy. The results showed that protoplumericin A and 13-O-p-coumaroylplumieride had high antifungal activities, which inhibited fungal growth by 72.97% and 69.92%, respectively. Scanning electron microscope observation revealed irregular mycelial shrinkage, surface collapse, or fuzzy symptoms. This study provides support for the further development of plant-source fungicides to control P. capsici. KeywordsAllemanda neriifolia; Phytophthora capsici; iridoids; antifungal activity 在天然药物化学成分的研究中,环烯醚萜类成分一直是研究的热点,亦是寻找和发现天然药物生物活性成分的重要来源。据不完全统计,至今从植物中发现的环烯醚萜类化合物有上千个。环烯醚萜类成分种类繁多、结构复杂,包括普通、裂环、二聚体等各种类型[1]。硬枝黄蝉(Allemanda neriifolia Hook.)是一种夹竹桃科黄蝉属植物,主要成分是环烯醚萜类化合物,有报道称其环烯醚萜类内酯成分黄蝉花定、鸡蛋花素等具有抗肿瘤和抗真菌的生物活性[2]。Kupchan等[3]从软枝黄蝉的根乙醇提取物中分离出黄蝉花定、黄蝉花辛和黄蝉花素,并发现其具有抗白血病的活性。Abe
2025年2月热带农业科学第45卷第2期 34 等[4]先后从黄蝉的茎、叶甲醇提取物中分离出21种环烯醚萜类化合物。有报道[5-7]称,从软枝黄蝉的根甲醇提取物分离出2个新的环烯醚萜糖苷类化合物plumieride coumarale和plumieride coumarate glucoside,且发现分离的鸡蛋花甙具有强烈的抗皮肤真菌的活性。Mohammed等[8]研究也表明,从夹竹桃科的6个属中鉴定出444种化合物,报道了11种生物活性,包括抗菌、抗炎等。辣椒疫霉菌Phytophthora capsici是一种毁灭性的卵菌病原体,可侵染许多重要的茄科和葫芦科等作物,造成重大经济损失[9]。苟亚峰等[10]研究发现,硬枝黄蝉无水乙醇提取物对辣椒疫霉菌P. capsici具有很强抑菌活性。本实验以P. capsici为供试病原菌,测定了从硬枝黄蝉中分离的4种环烯醚萜类化合物对P. capsici的抑菌活性及供试病原菌表观形态的影响,为进一步研制防治P. capsici的植物源杀菌剂提供依据。1 材料与方法1.1试材1.1.1 供试病原菌辣椒疫霉菌(P. capsici)由中国热带农业科学院香料饮料研究所植物保护研究室提供。1.1.2 供试化合物四种环烯醚萜类化合物plumieride 、protoplumericin A、13-dexoylplumieride和13-O-p-coumaroylplumieride,由中国热带农业科学院香料饮料研究所病虫害防控研究室采用硅胶柱层析等方法从硬枝黄蝉乙醇提取物中分离得到,并经过波谱学手段鉴定确定化合物结构和名称。1.2 方法1.2.1 抑菌活性测定参照方中达等[11]、慕立义[12]的生长速率法测定抑菌活性,具体方法:分别准确称取0.04 g供试的4种环烯醚萜类化合物,先用1 mL无水乙醇助溶,再用无菌蒸馏水定容至10 mL,配制成浓度为4 000 mg/L的母液;再用无菌蒸馏水稀释依次制成浓度为3 000、2 000 mg/L的药液;然后将培养基融化并冷却至50℃左右时,把1 mL不同浓度的4种环烯醚萜类化合物分别与9 mL/皿的PDA培养基混合,摇匀,待其凝固后,取菌落直径为5 mm、培养好的辣椒疫霉菌的外层菌块放置于培养皿中央,以加入等体积无水乙醇灭菌水的PDA培养平板为对照,每个处理3次重复;接种后,在人工气候培养箱(25~28℃)下培养5 d,采用十字交叉法分别测量菌落直径,求其平均值,抑菌率计算过程如下。菌落直径(cm)=测量菌落直径平均值–0.5 对照菌落直径–处理菌落直径抑菌率=对照菌落直径×100%1.2.2 化合物对辣椒疫霉菌菌丝表观形态的影响通过扫描电镜(SEM)观察4种环烯醚萜类化合物plumieride、protoplumericin A、13-dexoylplumieride和13-O-p-coumaroylplumieride处理的辣椒疫霉菌与对照菌丝的表观形态差异。具体方法:将空白对照和药剂处理组培养3~5 d的菌落采用点接触法在菌落边缘向下倾斜45°插入已灭菌盖玻片,于28℃继续培养3~5 d后取出载玻片,将带菌面朝下浸入5%戊二醛溶液中固定24 h,用PBS缓冲溶液(0.2 mol/L,pH 7.4)洗涤3次,15 min/次;依次用浓度为50%、70%、90%、100%的乙醇溶液进行2次洗脱,每次20 min;用醋酸异戊酯置换20 min,六甲基二硅胺烷浸泡30 min,风干,用导电胶固定到电台上,喷金约30 s,最后进行扫描电镜观察、拍照。2 结果与分析2.1 四种环烯醚萜类化合物对辣椒疫霉菌菌落生长的影响在供试浓度4 000 mg/L时,4种环烯醚萜类化合物plumieride、protoplumericin A、13-dexoylplumieride和13-O-p-coumaroylplumieride对辣椒疫霉菌菌落生长表现出不同程度的抑制作用,其中protoplumericin A和13-dexoylplumieride两种化合物处理后辣椒疫霉菌菌落较小,抑制活性较高,抑菌效果见图1。四种环烯醚萜类化合物plumieride、protoplumericin A、13-dexoylplumieride和13-O-pcoumaroylplumieride在供试浓度4 000、3 000、2 000 mg/L下对辣椒疫霉菌抑菌率见表1。从表1中可以看出,4种环烯醚萜类化合物在3种供试浓度下对辣椒疫霉菌抑菌率不同,其中,在供试浓度为4 000 mg/L时,protoplumericin A和13-O-p-coumaroylplumieride两种化合物对辣椒疫
苟亚峰等硬枝黄蝉中4种环烯醚萜类化合物对辣椒疫霉菌的抑菌活性研究- 35 -霉菌抑菌率均达到了60%以上,分别为69.92%和72.97%。plumieride和13-dexoylplumieride两种化合物对辣椒疫霉菌抑菌率较低,分别为30.06%和32.39%。A. CK;B. protoplumericin A;C. plumieride;D. 13-O-p-coumaroylplumieride;E. 13-dexoylplumieride。图1四种化合物对辣椒疫霉菌菌落生长的影响表1 四种环烯醚萜类化合物对辣椒疫霉菌菌丝生长抑菌率测定结果平均抑菌率/% 化合物种类4 000 mg·L-1 3 000 mg·L-1 2 000 mg·L-1plumieride 30.06 % 23.40 % 16.77 % 13-O-p-coumaroylplumieride 69.92 % 56.11 % 51.49 % protoplumericin A 72.97 % 59.27 % 52.33 % 13-dexoylplumieride 32.39 % 26.01 % 16.78 % 2.2 辣椒疫霉菌菌丝表观形态扫描电镜观察结果在2.1的基础上,通过扫描电镜就抑菌活性较好的2种环烯醚萜类化合物13-O-P-coumaroylplumieride和protoplumieride A对辣椒疫霉菌菌丝表观形态的影响进行观察、拍照,结果见图2。从图2可以看出,13-O-P-coumaroylplumieride和protoplumieride A两种化合物以供试浓度2 000 mg/L处理辣椒疫霉菌后,辣椒疫霉菌菌丝表观形态发生了变化。对照组(CK)菌丝表面光滑,而13-O-P-coumaroylplumieride和protoplumieride A处理组菌丝出现不规则收缩,菌丝表面塌陷或模糊等。图2两种化合物对辣椒疫霉菌表观形态的影响3 讨论与结论从植物天然产物中寻找结构新颖、对病虫害防治效果明显的化合物逐渐成为经济作物主要病虫害防治的重要途径。由天然产物研发的生物农药因具有易降解、低残留、高效等优势而被广泛应用,这不仅可减少病虫害对药剂产生抗药性,而且还能大大降低化学农药对环境的污染,这也符合海南自由贸易港建设中生态环境保护的需求。Posri等[13]从托叶黄檀茎中分离得到黄檀桐等活性化合物,其对Pythium insidiosum具有较高抑菌活性。植物精油在植物病原真菌防治中同样发挥着重要作用,如β-石竹烯、芳樟醇、柠檬烯、β-
2025年2月热带农业科学第45卷第2期 36 蒎烯、α-蒎烯、香芹酚、百里酚等[14]。Oliceira等[15]发现,从百里香叶片中提取的精油对镰刀菌Fusarium oxysporum、炭疽菌Colletotrichum gloeosporioides等具有抑菌活性,其中具抑菌活性的主要成分是香芹酚和百里酚。国外Khan等[16]测定大蒜根系分泌物对胡椒瘟病菌(由辣椒疫霉菌P. capsici侵染引起的病害)细胞形态的影响发现,大蒜分泌物对P. capsici菌丝生长具有明显抑制作用。Divya等[17]报道称,外源性应用茉莉酸和水杨酸可减轻辣椒疫霉的症状,水杨酸盐和茉莉酸盐防御作用的增强可能是控制疫病的一种途径。该研究结果与本研究结果一致,从硬枝黄蝉中分离的4种环烯醚萜类化合物中的protoplumericin A和13-O-p-coumaroylplumieride对P. capsici菌丝表观形态具有一定的影响,扫描电镜观察发现,该2种环烯醚萜类化合物处理后,P. capsici菌丝出现不规则收缩,菌丝表面呈现塌陷或模糊等症状,但抑菌活性与该2种环烯醚萜类化合物浓度之间的关系以及具体抑制P. capsici菌丝生长的作用靶标还需要进一步研究。参考文献[1] 廖铁松, 闵建新, 潘玲玲, 等. 茜草科植物环烯醚萜类化合物的研究进展[J]. 中草药, 2018, 49(6): 1 437-1 450. [2] Pai B R, Subramaniam P, Rao U. Isolation of plumericin and isoplumericin from Aliamanda cathartica Linn[J]. Indian J Chem, 1970, 8(9): 851. [3] Kupchan S M, Dessertine A L, Blaylock B T, et al. Isolation and structural elucidation of allamandin, an antileukemic iridoid lactone from Allamanda cathartica[J]. The Journal of Organic Chemistry, 1974, 39(17): 2 477-2 482. [4] Abe F, Mori T, Yamauchi T. Iridoids of Apocynaceae. III. minor iridoids from Allamandaneriifolia[J]. Chemical and Pharmaceutical Bulletin, 1984, 32(8): 2 947-2 956. [5] Coppen J J W. Iridoids with algicidal properties from Allamanda cathartica[J]. Phytochemistry, 1983, 22(1): 179- 182. [6] Coppen J J W, Cobb A L. The occurrence of iridoids in Plumeria and Allamanda[J]. Phytochemistry, 1983, 22(1): 125-128. [7] Tiwari T N, Pandey V B, Dubey N K. Plumieride from Allamanda cathartica as an antidermatophytic agent[J]. Phytotherapy Research, 2002, 16(4): 393-394. [8] Mohammed, A E, Abdul-Hameed Z H,, Alotaibi M O, et al. Chemical diversity and bioactivities of monoterpene indole alkaloids (MIAs) from six apocynaceae genera[J]. Molecules, 2021, 26(2): 488. [9] Hudson O, Waliullah S, Hand J, et al. Detection of Phytophthora capsici in irrigation water using loop-mediated isothermal amplification[J]. Journal of Visualized Experiments, 2020: (160). [10] 苟亚峰, 胡林峰, 曾煊, 等. 硬枝黄蝉乙醇提取物中抑制胡椒瘟病菌活性成分研究[J]. 植物保护, 2020, 46(6): 183-185+193. [11] 方中达. 植病研究方法(3版)[M]. 北京: 中国农业出版社, 1998. [12] 慕立义. 植物化学保护研究方法[M]. 北京: 中国农业出版社, 1994: 79-81. [13] Posri P, Suthiwong J, Thongsri Y, et al. Antifungal activity of compounds from the stems of Dalbergia stipulaceaagainst Pythium insidiosum[J]. Natural Product Research, 2021, 35(17): 2 823-2 830. [14] Karpiński T M. Essential oils of Lamiaceae family plants as antifungals[J]. Biomolecules, 2020, 10(1): 103. [15] Oliveira T S, Silva-Filho C S, Malveira E A, et al. Antifungal and antibiofilm activities of the essential oil of leaves from Lippia gracilis Schauer against phytopathogenic fungi[J]. Journal of Applied Microbiology, 2021, 130(4): 1 117-1 129. [16] Khan M A, Cheng Z H. Influence of garlic root exudates on cyto-morphological alteration of the hyphae of Phytophthora capsici, the cause of Phytophthora blight in pepper[J]. Pakistan Journal of Botany, 2010, 42(6): 4 353- 4 361. [17] Divya K, Artur P, Sweda S, et al. Non-targeted metabolite profiling reveals host metabolomic reprogramming during the interaction of black pepper with Phytophthora capsici[J]. International Journal of Molecular Sciences, 2021, 22(21): 11 433. (责任编辑林海妹)
2025年2月热带农业科学第45卷第2期Feb. 2025 CHINESE JOURNAL OF TROPICAL AGRICULTURE Vol.45, No.2 收稿日期 2024-01-25;修回日期 2024-03-18 基金项目 三亚市科技创新专项项目“优异抗虫豇豆种质资源鉴定、评价及应用”(No. 2022KJCX19);海南省重点研发计划“豇豆内禀抗豆大蓟马次生代谢物发掘及其生物合成路径解析”(No. ZDYF2021XDNY302);海南省自然科学基金“豇豆挥发物DMNT参与的豇豆内禀抗虫模型及其生物合成调控路径解析”(No. 323RC521)。第一作者何云川(1995—),男,博士研究生,研究方向为农业昆虫与害虫绿色防治,E-mail:heyunchuan@zju.edu.cn。通信作者 周瀛(1986—)女,博士,研究员,研究方向为农业昆虫与害虫绿色防治,E-mail:yzhyzb@zju.edu.cn。豇豆种质资源对普通大蓟马的抗性鉴定技术规程何云川1,2张想芬1,2周瀛1祝增荣1,2(1. 浙江大学海南研究院海南三亚 572025;2. 浙江大学昆虫科学研究所浙江杭州 310058)摘要普通大蓟马是豇豆上危害最为严重的害虫之一,严重影响了豇豆的产量与品质。为了经济、有效、绿色防控田间有害生物对作物的危害,抗性品种选育成为了一种重要的防治措施。抗虫种质资源是培育抗虫豇豆品种的重要遗传资源基础,本文基于文献报道和实验研究总结出豇豆种质资源对普通大蓟马抗性的鉴定技术规程,规程内容主要包括:规程的适用范围、规范性文件引用、术语和定义、室内和田间鉴定方法。该规程将为豇豆种质资源对普通大蓟马抗性的鉴定标准制定提供重要参考,也对豇豆抗虫新品种的选育具有重要意义。关键词豇豆;种质资源;普通大蓟马;抗性;技术规程中图分类号S436.43 文献标识码 A DOI: 10.12008/j.issn.1009-2196.2025.02.007 Technical Procedure for the Assessment of Cowpea Germplasm Resistance to Thrips (Megalurothrips usitatus) HE Yunchuan1,2 ZHANG Xiangfen1,2 ZHOU Ying1 ZHU Zengrong1,2(1. Hainan Institute, Zhejiang University, Sanya, Hainan 572025, China; 2. Institute of Insect Science, Zhejiang University Hangzhou, Zhejiang 310058, China) Abstract Megalurothrips usitatus is one of the most serious cowpea pests and severely affects yield and quality. To control the damage caused by pests to crops economically, effectively, and environmentally friendly, the breeding of resistant varieties has become an important control measure. Insect-resistant germplasm resources constitute a vital genetic basis for cultivating insect-resistant cowpea varieties. Based on literature reports and experimental studies, this paper summarized the technical procedure for evaluating resistance to M. usitatus in cowpeas, which mainly includes the scope of procedure application, reference of normative documents, terms and definitions, and indoor and field identification methods. This technical procedure will provide a crucial reference for the formulation of evaluation standards for thrips resistance in cowpea germplasms and is highly important for the breeding of new insect-resistant cowpea varieties. Keywords cowpea; germplasm; Megalurothrips usitatus; resistance; technical procedure 豇豆是重要的经济作物,也是农民增收、农业增效的重要来源之一。豇豆在我国种植总面积超过33万亩(1亩≈667 m2),也是海南省重要的冬种瓜菜之一[1-2]。近年来,由于豇豆田间种植结构单一,生态系统生物多样性降低等原因,导致普通大蓟马在海南种植区危害尤为严重,呈现出连年爆发危害的现状,严重威胁着海南豇豆种植业的发展。普通大蓟马主要危害豇豆的嫩叶、花和豆荚,造成豇豆生长点发育受阻,叶片卷曲,花苞发育不正常,豆荚生长畸形,且两端呈现黑头黑尾等症状,严重影响豇豆的产量和品质[3-5]。此外,为防治普通大蓟马的猖獗发生,常年采用化学农药防治而带来的一系列不良影响已日趋严重,如普通大蓟马抗药性增强、豇豆农残超标以及田间生态系统生物多样性降低等[6-9]。这些问题严重制约了豇豆种植业的可持续发展。因此,建立高效的豇豆有害生物绿色防治体系是亟需解决的科学问题。大量研究实践表明,选育和利用抗虫品种是防治虫害最为安全、经济、有效的措施之一,有利于农业可持续发展和保护人类自身生存环境。目前,许多国家均已将培育抗虫品种作为国家育
2025年2月热带农业科学第45卷第2期38 种的重要目标之一。在我国,开展对农作物的抗病虫性鉴定已有相当长的历史,在国家及各省科技项目的资助下,科技工作者开展了包括水稻、小麦、玉米等主粮作物;棉花、大豆等重要经济作物的种质资源对主要病虫害抗性的鉴定和评价工作,筛选出了一批优异的抗性种质资源,为抗性育种提供了宝贵的种质资源[10-13]。其他农作物的种质资源抗病虫性鉴定进展与主粮作物及重要经济作物相比较为缓慢,而豇豆种质资源的病虫害抗性评价工作及抗性种质资源利用在国内目前尚未系统开展,国内相关报道仅在20世纪90年代有部分的抗蚜虫和抗锈病研究[14-15],尚无主要害虫普通大蓟马的抗性评价工作。因此,亟需快速推进此方面的相关研究,以加强对现有抗虫种质优良性状的挖掘与利用,加快种子的研发创新。基于以上背景,本文总结了豇豆种质资源对普通大蓟马抗性的鉴定技术规程,以期为制定豇豆种质资源对普通大蓟马抗性的鉴定标准提供参考。1规程范围本标准规定了普通大蓟马抗性鉴定技术方法和抗性评价方法。本标准适用于豇豆品种、中间材料及种质资源等对普通大蓟马的抗性鉴定。2术语和定义下列术语和定义适用于本文件。2.1抗性鉴定通过适宜技术方法鉴别农作物对其特定害虫的抵抗水平。2.2虫量比值法虫量比值法是一种根据所求值进行抗性等级划分的抗性评价方法,即某参试品种的蓟马量均值除以全部参试品种的蓟马量均值。3生物学特性3.1普通大蓟马普通大蓟马(Megalurothrips usitatus Bagnall)又名豆花蓟马、豆大蓟马,隶属缨翅目(Thysanotpera)锥尾亚目(Terebrantia)蓟马科(Thripoidae)大蓟马属(Megalurothrips),是豇豆上的重要害虫之一。3.2形态特征普通大蓟马属不完全变态发育,历经卵、若虫、伪蛹、成虫4个阶段,各个阶段形态特征如下。卵:卵粒肾型长约 0.2 mm(图1-A)。1龄若虫:初孵1龄若虫白色透明,体长约0.52 mm,眼点红色,活动性强(图1-B)。2龄若虫:2龄若虫体长0.98 mm,体色渐黄,且随着体色向橙红色的转变,开始进入预蛹期(图1-C)。预蛹:预蛹期具白色透明的短翅芽,触角向前。伪蛹:蛹期翅芽伸达腹部2/3处、触角向后(图1-D)。成虫:雌成虫体长约1.6 mm,棕色至暗棕色(图1-E),雄成虫体色与雌成虫相似,但体型略小(图1-F)。触角念珠状略向前延伸,具复眼和单眼,口器锉吸式,狭窄的翅周缘着生细长缨毛。A.卵;B.一龄若虫;C.二龄若虫;D.伪蛹期;E.雌成虫;F.雄成虫。图1普通大蓟马不同发育阶段示意图
何云川等豇豆种质资源对普通大蓟马的抗性鉴定技术规程-39 -3.3为害特点普通大蓟马为害豇豆整个生育期,几十或几百头普通大蓟马通过聚集取食嫩叶(图2-A)、花(图2-B)和豆荚(图2-C),造成嫩叶和花芽卷曲、坏死,进而影响光合作用;豇豆花过早脱落,影响豆荚的发育形成;豆荚黑头黑尾症状,影响豇豆品质和经济价值。普通大蓟马还能传播Tobacco streak virus (TSV)病毒病为害豇豆植株[16],导致豇豆植株大规模死亡进而造成严重减产,对豇豆产业的健康发展造成了严重威胁。A. 普通大蓟马为害造成嫩叶卷曲;B. 普通大蓟马为害花;C. 普通大蓟马为害豆荚,形成黑头黑尾症状。图2豇豆植株被普通大蓟马为害状况3.4生活习性与发生规律普通大蓟马成虫活跃、善飞、惧光,雌成虫主要为孤雌生殖,也偶有两性生殖;卵散产于叶肉组织内,每雌产卵22~35粒,若虫发育到3龄末期停止取食,坠落在表土。普通大蓟马在海南、广西、贵州一年发生17~18代,世代重叠,终年繁殖,主要危害豇豆。在豇豆开花后,普通大蓟马的种群数量迅速增加,出现一段时间的为害高峰,并持续为害到豇豆生长结束[17]。由于海南南部豇豆普遍在冬季种植(春节前后上市),为普通大蓟马提供了大量的寄主。同时海南南部冬季气候干燥少雨,温度适宜。因此,普通大蓟马在海南南部冬季田间发生严重[17]。4室内抗性评价4.1虫源准备田间采集普通大蓟马成虫,在温度(26±0.5)℃,湿度(60±5)%,光周期14∶10 (L∶D) h的光照培养箱内用豇豆豆荚继代饲养备用。4.2鉴定材料准备将参试豇豆材料种植在隔离区(无虫源植物房),每个处理3株,每株10朵花,重复3次。4.3接虫在豇豆开花期,将2龄老熟普通大蓟马若虫接在豇豆花上,每朵豇豆花接虫20头。4.4接虫后的管理豇豆开花期接虫后,罩上120目尼龙网罩防止普通大蓟马逃逸,其余管理常规进行。5田间抗性评价5.1田块选择豇豆田选在普通大蓟马重发区,有浇水条件、光照充足、地势平坦、土壤肥沃的田块。5.2试验设计每份参试豇豆材料每个小区种植6穴,每穴之间的间距为20 cm,每垄间的行距为90 cm,重复5次,试验小区按照完全随机试验设计。待出苗后,每份参试豇豆材料每小区各6株,5次重复共计30株。种植时间与当地大田生产种植时间相同。5.3田间管理豇豆整个生育期内不施用任何农药,其余农事操作常规管理。6室内与田间花期抗性评价方法普通大蓟马盛发期主要危害豇豆花(图3),故以花作为豇豆种质资源对普通大蓟马抗性评价的主要评价指标。6.1室内抗性评价方法豇豆花接虫5 d后,根据Jackai 等[18]的方法,对豇豆花进行受害分级(表1)。统计各品种的为害级别,并按式(1)计算其为害指数[19]。花受害指数法的抗性分级标准以花受害指数最高与最低差值等为基准划分为高抗(HR)、抗(R)、中抗(MR)、感(S)、高感(HS)5个级别[19]。
2025年2月热带农业科学第45卷第2期40 图3普通大蓟马取食危害豇豆花症状结构图表1普通大蓟马为害豇豆花的受害分级标准花受害分级花危害症状1 豇豆花旗瓣无蓟马危害状或变色3 豇豆花旗瓣外侧无明显蓟马危害状或变色5 豇豆花旗瓣、翼瓣均出现明显的蓟马危害状或变色7 豇豆花旗瓣、翼瓣和龙骨瓣蓟马危害状严重,有明显取食点,造成花瓣色泽变褐,木质化9 豇豆花旗瓣、翼瓣和龙骨瓣蓟马危害状严重,有明显取食点,花瓣严重褐色化,花托和花瓣干燥皱缩( ) =n giL gL LIN M ······························· (1)式中:Ii ,受害指数,单位为百分率(%);Ln,各参试豇豆材料受害花数,Ln=1,2,3,…,30;Lg,受害花分级值,Lg=1,3,5,7,9;NL,各参试豇豆材料调查总花数,NL=30;Mg,各参试豇豆材料受害花分级最大值,Mg=9。6.2田间抗性评价方法在豇豆开花期(小区内50%的植株见花的日期)后的一周进行豇豆花的采集,对每个小区的每个参试豇豆材料的每个重复随机采取10朵花,每个参试豇豆材料共收集50朵花。将每个材料的单朵花放入PE塑料自封袋(7 cm×9 cm),并加入75%的酒精保存,带回室内光学显微镜下进行鉴定计数,记录每朵花普通大蓟马成虫和若虫数。根据式(2)计算普通大蓟马的虫量比值确定豇豆花被普通大蓟马为害的抗性分级标准[20-21],划分为高抗(HR)、抗(R)、中抗(MR)、感(S)、高感(HS)5个级别(表2)。虫量比值按式(2)计算:+ =N AffMMIN ······························· (2)式中:If,虫量比值,单位为百分率(%);MN,各参试豇豆材料单朵花普通大蓟马若虫数;MA,各参试豇豆材料单朵花普通大蓟马成虫数;Nf,全部参试豇豆材料单朵花普通大蓟马的平均虫数。
何云川等豇豆种质资源对普通大蓟马的抗性鉴定技术规程-41 -表2普通大蓟马为害豇豆花的抗性分级标准抗性等级标准抗性类型1 If<M-1.5S HR 3 M-1.5S≤If <M-0.5SR 5 M-0.5S≤If <M+0.5SMR 7 M+0.5S≤If <M+1.5SS 9 ≥M+1.5SHS 注:M. 全部参试豇豆材料的加权平均数的总平均数;S. 全部参试豇豆材料的加权平均数的标准差。7接虫后与鉴定后材料处理室内接虫后剩余的普通大蓟马、鉴定后的虫害材料作杀虫处理。8结果判定8.1有效性判定当田间全部参试豇豆材料的M-1.5*S值不为负值时,鉴定结果有效。8.2抗性级别判定当室内抗性评价结果与田间抗性评价结果不一致时,以抗性较弱的级别作为该材料的最终抗虫性级别。9鉴定与评价结果记载鉴定与评价结果记录参见表3。表3普通大蓟马抗性鉴定与评价记录表编号参试材料材料来源室内抗性评价结果田间抗性评价结果综合抗性评价结果鉴定地点:豇豆田类型:海拔高度:经度:纬度:播种日期:接虫日期:开花日期:调查日期:调查人:参考文献[1]李甜瑞, 方贯娜, 万秀娟, 等. 我国长豇豆品种选育研究进展[J]. 北方园艺, 2023(11): 129-135. [2]孙铁玉海南冬季瓜菜产销现状与发展对策[J]. 中国瓜菜, 2022, 35(7):105-109. [3]He Y C, Gao Y, Hong H N, et al. Megalurothrips usitatusdirectly causes the black-heads and black-tail symptoms of cowpea along with the production of insect-resistance flavonoids[J]. Plants, 2023, 12(22): 3 865. [4]吴圣勇, 谢文, 刘万才, 等. 我国豇豆蓟马研究进展及综合防控措施[J]. 植物保护, 2021, 50(2): 10-18. [5]潘雪莲, 杨磊, 金海峰, 等. 豆大蓟马在海南发生与防治的研究进展[J]. 热带生物学报, 2021, 12(4): 508-513. [6]邱海燕, 付步礼, 谭魁孙, 等. 海南豆大蓟马田间种群对多种杀虫剂的抗性监测[J]. 中国植保导刊, 2022, 42(11): 67-71. [7]袁琳琳, 潘雪莲, 陆容材, 等. 海南普通大蓟马对拟除虫菊酯药剂抗性监测及钠通道突变检测[J]. 植物保护, 2023, 49(3): 60-67. [8]金国藩, 蒋士强, 王静, 等. 从“海南毒豇豆”事件看农产品食品安全监管与农残检测[J]. 食品安全导刊, 2010,(4): 23-25. [9]杨普云, 李萍, 任彬元, 等. 我国农作物病虫害化学防控技术的环境成本分析[J]. 中国植保导刊, 2019, 39(6): 27- 30. [10]卓乐, 邓永阳, 田琴琴, 等. 南方双季稻品种对白背飞虱的抗性鉴定[J/OL]. 分子植物育种, (2023-06- 30)[2024-01-03] https://kns.cnki.net/kcms2/detail/46.1068.S. 20230629.1855.008.html. [11]Chiriboga Morales X, Tamiru A, Sobhy IS, et al. Evaluation of african maize cultivars for resistance to fall armyworm
2025年2月热带农业科学第45卷第2期42 Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) larvae [J]. Plants, 2021, 10(2): 392. [12]Zhang L J, Geng M M, Zhang Z, et al. Molecular mapping of major QTL conferring resistance to orange wheat blossom midge (Sitodiplosis mosellana) in Chinese wheat varieties with selective populations[J]. Theoretical and Applied Genetics, 2020, 133: 491-502. [13]Muhammad R, Saifullah A, Usman A M, et al. Evaluation of cotton germplasm for morphological and biochemical host plant resistance traits against sucking insect pests complex [J]. Journal of Cotton Research, 2021, 4: 18. [14]韩粉霞, 刘京宝, 冯庆平. 豇豆优异种质综合评价[J]. 植物遗传资源科学, 2001, 2: 34-37. [15]王佩芝, 冯庆平, 阎家廉, 等. 豇豆优异种质资源综合评价[J]. 作物品种资源, 1995, 5: 14-16. [16]Sonali Shukla S, Kalyani G, Kulkarni N, et al. Mechanism of transmission of tobacco streak virus by Scirtothrips dorsalis, Frankliniella schultzei and Megalurothrips usittusin groundnut, Arachis hypogaea L[J]. Journal of Oilseeds Research, 2005, 22(1): 215-217. [17]唐良德, 梁延坡, 韩云, 等. 海南豇豆蓟马发生为害调查及蓝板监测技术研究[J]. 中国植保导刊, 2015, 35: 53-57. [18]Jackai L E N, Singh S R. Screening techniques for host plant resistance to insect pest of cowpea[J]. Crop Grain Legume Bulletin, 1988(35): 2-18. [19]郑逆, 李庆, 蒋春先, 等. 辣椒品种对侧多食跗线螨的抗性鉴定方法研究[J]. 西南师范大学学报(自然科学版), 2010, 35 (5): 91-94. [20]张中润, 高燕, 黄建峰, 等. 不同芒果品种对蓟马的抗虫性评价[J]. 南方农业学报, 2020, 51: 1 591-1 597. [21]盖钧镒,崔章林. 大豆抗食叶性害虫育种的鉴定方法与标准[J]. 作物学报, 1997(4): 400-407. (责任编辑龙娅丽)
2025年2月热带农业科学第45卷第2期Feb. 2025 CHINESE JOURNAL OF TROPICAL AGRICULTURE Vol.45, No.2 收稿日期 2024-04-08;修回日期 2024-05-15 基金项目 江西重点研发计划(No.20223BBF61015);海南省自然科学基金(No.321RC618);武汉市卫生健康委科研基金资助(No.WG20D11)。第一作者李得铭(1993—),男,硕士研究生,助理农艺师,研究方向为土传病害防治,E-mail:83211989@qq.com。通信作者 黄俊生(1963—),男,博士,研究员,研究方向为微生物资源与利用,E-mail:h888111@126.com。绿农林®41号复合微生物菌肥对番茄生长影响及对青枯病防治作用李得铭1汪军1翟子翔1 邓涛2曹智醇3黄俊生1(1. 中国热带农业科学院环境与植物保护研究所海南海口 571101;2. 江西省科学院微生物研究所江西南昌 330095;3. 长江航运总医院湖北武汉 430000)摘要为明确绿农林®41号复合微生物菌肥对田间番茄生长和青枯病防治影响,在两地番茄生长过程中施用复合微生物菌肥,测定根系土壤养分含量、植株生物学性状及青枯病发病率。结果显示,与对照相比,施用复合微生物菌肥处理后土壤有机质增长11.5%~18.2%,速效钾、速效磷、速效氮分别增长19.3%~49.0%、21.6%~34.0%、7.0%~2.8%;植株株高、茎围和叶面积增加14.0%~33.5%、25%~46%、24.0%~37.0%,叶绿素和净光合作用的增幅分别为17.2%~17.8%和5.7%~6.0%。番茄青枯病的发病率下降24.0%~32.0%,果实水溶性总糖和单粒果实重量分别增加1.5%~2.7%、8.4%~11.6%。表明复合微生物菌肥在改良土壤、促进番茄生长、防治番茄青枯病方面具有显著作用。关键词微生物菌肥;番茄;生长;青枯病中图分类号S641.2 文献标识码 A DOI: 10.12008/j.issn.1009-2196.2025.02.008 Lvnong lin® 41 Compound Microbial Fertilizer on Tomato Growth and Control of Ralstonia solanacearumLI Deming1 WANG Jun1 ZHAI Zixiang1 DENG Tao2 CAO Zhichun3 HUANG Junsheng1(1. Institute of Environment and Plant Protection, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan 571101, China; 2. Institute of Microbiology, Jiangxi Academy of Sciences, Nanchang, Jiangxi 330095, China; 3. General Hospital of the Yangtze-River Shipping, Wuhan, Hubei 430000, China) Abstract To clarify the impact of Lvonglin ® 41 compound microbial fertilizer on tomato growth and its ability to control the effect on tomato bacterial wilt caused by bacterial wilt in the field, the compound microbial fertilizer was applied during thegrowth of tomatoes at two locations to determine the nutrient content of root-soil, plant biological characteristics and incidence of bacterial wilt. The results showed that compared with the control, soil organic matter increased by 11.5%-18.2% after the application of compound microbial fertilizer, and available potassium, available phosphorus, and available nitrogen increased by 19.3%-49.0%, 21.6%-34.0%, and 7.0%-2.8% respectively; plant height, stem circumference, and leaf area increased by 14.0%-33.5%, 25%-46%, and 24.0%-37.0%, and chlorophyll and net photosynthesis increased by 17.2%-17.8% and 5.7%-6.0% respectively. The incidence of tomato bacterial wilt decreased by 24.0%-32.0%, and the total water-soluble sugars in fruits and the weight of individual fruits increased by 1.5%-2.7%, and 8.4%-11.6%, respectively. The results showed that thecompound microbial fertilizer had significant effects on improving soil quality, promoting tomato growth, and controlling tomato bacterial wilt. Keywords microbial fertilizer; tomato; growth; bacterial wilt 我国华南地区土壤类型多数为砖红壤,在番茄种植过程中,由于常年施用化肥使得土壤板结、微生物群落和土壤理化性质失衡,极易导致田间土传病害的发生[1-4]。微生物菌肥作为重要的绿色防控手段,在改善土壤结构、增加土壤的肥力、提高植物的抗病性等方面虽已得到广泛认可,但由于土壤类型、气候和环境条件多样[5-10],同时微生物菌肥的施用方式和产品生产工艺也往往影响着其功能效果的发挥[11]。因此,为进一步明确微生物菌肥对番茄青枯病的防控效果[12-17],本实
2025年2月热带农业科学第45卷第2期 44 验在番茄种植过程中施用由中国热带农业科学院环境与植物保护研究所研制的绿农林®41号复合微生物菌肥,探讨其对土壤理化性质和番茄产量及番茄青枯病防控效果的影响,为该微生物菌肥今后在番茄生产上的大面积推广应用提供支撑。1 材料与方法1.1材料供试菌肥:绿农林®41号复合微生物菌肥,由中国热带农业科学院环境与植物保护研究所微生物资源研究与利用实验室研制,由具有发明专利的复合菌群枯草芽孢杆菌、解淀粉芽孢杆菌、绿色木霉、淡紫拟青霉组成,有效孢子含量≥50亿/g,有机质≥40%,总养分(N+P2O5+K2O)≥12%。试验时间:2019年10月14日至2019年12月17日。试验开始前耕层土壤基本理化性质,1号实验地:速效氮40.93 mg/kg、速效磷106.7 mg/kg、速效钾127.5 mg/kg、有机质11.21 g/kg。2号实验地:速效氮46.84 mg/kg、速效磷110.9 mg/kg、速效钾256.7 mg/kg、有机质13.95 g/kg;1.2 方法1.2.1 试验设计地点:海南省琼海市桥头桥村番茄采摘基地(E: 110°50′64.88′、′N19°22′62.22′′),地处热带季风气候区北缘,光照充足,高温多雨,四季不明显,旱季和雨季分明。年均降雨量2 042.6 mm,年均气温24℃。年均日照数为2 155 h,占可照时数的49%。土壤以砖红壤为主,该土种土体深厚,质地多为砂质壤土。设置2个不同地块示范处理,采用小区排列对比方法。1、2号实验地:于2019年10月14日定植,品种为海南千禧番茄。种植行距60 cm、株距40 cm,共4行,2个相邻行为一组处理,每个处理50株,共100株。均统一管理。1号试验地设置1个对照组和1个处理组:1-1对照组,常规施肥+施用等体积清水;1-2处理组,常规施肥+绿农林®41号复合微生物菌肥。2号试验地设置1个对照组和1个处理组:2-1对照组,常规施肥+施用等体积清水;2-2处理组,常规施肥+绿农林®41号复合微生物菌肥。两地均在常规施肥的基础上,在番茄定植3周后,将绿农林®41号复合微生物菌肥稀释至500倍,浇灌于番茄根部;之后每隔3周按同样方法施用1次,共施4次。1.2.2项目调查两地实验同时进行,定植后全部统一管理,根据实验方案实施,每次施用微生物菌肥前,采集根系土壤测定土壤有机质、土壤基本性质。在每组实验组内随机选取10株,对植株生长状态、叶片生长状态、净光合作用、叶绿素、发病率、品质和产量等进行统计,实验数据的收集从第6周至第15周(大面积果实成熟期)。(1)土壤养分测定实验地第1次施肥后,每3周采集1次土壤,对两块番茄地土壤有机质进行测定,将实验土壤样品去除杂质,风干、过筛,运用灼烧法测量土壤有机质[20-21],采用HM-TYA型土壤肥料养分速测仪,对速效氮、速效磷、速效钾进行测定[22-25]。(2)土壤青枯病三重PCR检测从2个实验地中分别随机选择15株已发病番茄,利用CTAB缓冲液提取番茄根茎DNA粗提液直接进行青枯病三重PCR检测[26-28]。(3)净光合速率测定在晴朗的天气下测定,大气CO2浓度保持在(380±6.0)μmol/mol、大气温度26℃、相对湿度(60±4.0)%。测定不同土壤水分下光合作用的光响应过程,每组随机选取10株,分别从每株上、下部分选取3只健康叶片,每个叶片重复3次记录。采用LCPRO-SD便携式光合测定仪测定,时间为120 s,取平均值进行分析[29-30]。(4)叶绿素含量测定在自然光下每组随机选取10株,分别从每株上、下部选取3片健康叶片,每个叶片重复3次记录,用SY-S20手持式叶绿素检测仪测定[31-32]。(5)叶片生长状态测定每组随机选取10株,从每株上、下部分选取5只健康完整无虫眼植株的叶片,用YMJ-CH型智能叶面积测定系统定仪测定[33]。(6)番茄发病率、品质和产量测定大面积果实成熟(15周)后,每组随机选取10株番茄的第1、2穗果称重。同时用游标卡尺对果实长度、宽度进行测量,利用具有温度自动补偿功能的MSDR-P1高精度数显糖度计进行水溶性总糖测定[34-35]。1.2.3 数据处理采用Excel 2017对数据进行整理分析[18-19]。
李得铭等绿农林®41号复合微生物菌肥对番茄生长影响及对青枯病防治作用- 45 -2结果与分析2.1 不同处理对番茄种植地土壤养分的影响如图1所示,不同测定周期,1、2号实验地对照组有机质含量无明显变化,施用微生物菌肥后土壤有机质的含量均有明显提高。1号实验地,处理组与对照组相比土壤有机质增长率达18.2%;2号实验地,处理组与对照组相比土壤有机质增长率达11.5%。1-1. 1号实验地对照组;1-2. 1号实验地处理组;2-1. 2号实验地对照组;2-2. 2号实验地处理组。下同。图1不同处理对番茄种植地土壤有机质的影响种植15周时,番茄出现大面积果实成熟。如图2所示,此时番茄种植地土壤速效氮、速效磷、速效钾,实验组相比对照组均有不同程度的增加。1号实验地,处理组与对照组相比,速效钾、速效磷、速效氮分别增长49.0%、34.0%、7.0%。2号实验地,处理组与对照组相比,速效钾、速效磷、速效氮分别增长19.35%、21.6%、2.8%。图2不同处理对番茄种植地土壤基本性质的影响2.2 不同处理对番茄生长状态的影响两块番茄种植区在不同处理下植株整体生长状况存在明显差异。如图3所示,施用微生物菌肥的实验组整体生长速率高于对照组。1号实验地植株生长高度(图3-A),处理组比对照组高46.0%,植株茎围粗37.0%(图3-B)。2号实验地植株生长高度,处理组比对照组高25.0%,植株茎围粗24.0%。A. 番茄株高生长差异;B.番茄茎围生长差异。图3 不同处理对番茄生长状态的影响2.3 不同处理对番茄叶片生长状态的影响叶片作为主要器官之一,具有光合、呼吸、蒸腾等作用,在一定程度上,叶片发育情况可作为植株生长发育的衡量标准。如图4所示,两地图4不同处理对两地块番茄番茄叶面积生长差异
2025年2月热带农业科学第45卷第2期 46 块中处理组与对照组间叶片发育情况存在差异,1号实验地番茄叶面积相差14.0%,2号实验地番茄叶面积相差33.5%。2.4 不同处理对番茄叶片叶绿素和净光合作用的影响叶片叶绿素含量与光合速率、作物营养状况密切相关,生产上也往往依据叶色变化作为看苗诊断和肥水管理的重要依据。如图5所示,两地块番茄叶片发育过程中,对照组叶绿素含量(SPAD)变化幅度较小,处理组有小幅度提升。1、2号实验地叶绿素含量分别提升17.2%和17.8%。图5不同处理对番茄叶片叶绿素含量的影响植物生长过程中90%左右的干物质是由叶片进行光合作用转化而来,光合作用情况直接影响植株生长发育。如图6所示,两地块处理组在9周时净光合速率增幅最快,随后变化幅度较小。对照组间净光合速率增幅较为平缓。1、2实验地间净光合速率,实验组与对照组相比分别增加6.0%和5.7%。图6不同处理对番茄叶片净光合作用的影响2.5 不同处理对番茄发病率的影响在田间连作的情况下很容易造成番茄青枯病的爆发,两块番茄实验地均出现大量病株,其发病症状与青枯病类似,首先,顶部叶片萎蔫并下垂,然后下部叶片萎蔫,最后中间叶片萎蔫,也有一些单独一侧先萎蔫以及一整株叶片同时萎蔫、病茎表皮粗糙、茎中下部增生不定根或不定芽[36-38]。横切病茎,维管束变为褐色,挤压后切面可溢出白色菌液,以上可作为田间番茄青枯病诊断的重要凭证。在2个样地中,共检测了30株患病植物,如图7所示,结果均为阳性,结合两地田间发病症状,确定该病害为番茄青枯病。对两地块间番茄青枯病发病情况进行统计。番茄种植15周后发病,1号实验地中,1-1对照组发病率为32.0%,施用微生物菌肥的1-2处理组发病率仅为8.0%,同期相比发病率下降24.0%。2号实验地中,2-1对照组发病率为42.0%,施用微生物菌肥的2-2处理组发病率仅为10.0%,同期相比发病率下降32%。结果表明,施用微生物菌肥15周平均发病率为9.0%,对田间番茄青枯病的发生具有良好防控作用。A. I号地块发病植株检测结果;B. II号地块发病植株检测结果。M. DL 2 000 DNA Marker;1. 阴性对照;2~16. 田间发病植株。图7 田间病株的三重PCR检测2.6 不同处理对番茄品质和产量的影响在番茄生长发育过程中,田间病虫害、肥水供应状况、光照条件等可直接影响番茄品质和产量。不同处理可对番茄果实形态、果实品质及产量产生明显影响。处理组和对照组相比,整体果实更圆润,果实水溶性总糖含量1号实验地最高,

