










































3 讨论与结论
近年来研究表明,复合微生物菌肥在改良土壤性质、防治土传病害、提高产量和品质方面发挥重要作用。邵雪凤[39]、郭立佳等[40研究发现,氨基酸水溶肥可促进芽孢杆菌在土壤中的生长繁殖,使之分泌抑菌活性物质和促生长激素,增强植物对病原菌的防御能力。张洁梅等4筛选出对烟草青枯病有效防治的生防菌,该菌还能提高烟叶品质和产量。施用复合芽孢杆菌肥可以促进水稻生长,显著改善土壤理化性质,促使水稻植株产生抗病性,降低发病率[4]。本研究发现,田间番茄青枯病的发生,对照组与处理组相比发病率下降。
微生物菌肥对增强光合作用、促进营养物质吸收也有显著效果。有研究[43-44]表明,含有解淀粉芽孢杆菌的微生物菌肥可以促进水稻植株生长,并对水稻稻瘟病具有防治作用。施用微生物菌肥使黄瓜植株的光合指标值、光合作用显著增加,极大地促进黄瓜植株的生长[45]。汪军等[46]、罗静静等7指出,复合微生物菌肥比单一微生物菌肥的防病促生效果显著。本实验采用的“绿农林 {}^{registered}41 号”复合微生物菌肥中含有大量的复合芽孢杆菌,促进植物生长效果显著,这与邹成等的研究结果类似。本研究通过田间试验对比,验证了施用绿农林 ^{B}41 号复合微生物菌肥可明显提高番茄株高、茎围和叶面积,分别增加 25% 、 24% 和 33.5% ,同时叶绿素和净光合作用分别增加了17.8% 和 5.7% 。
微生物菌肥在提高作物产量、改善作物品质方面的作用也早已被证实。王丽丽等[48研究证实,微生物菌肥可显著提高番茄的VC和可溶性糖含量,从而改善番茄品质和产量。本研究结果表明,各处理果实整体更圆润,1号地和2号地水溶性总糖平均值比同组对照组高 2.7% ,产量高8.4% ,在改善番茄品质上具有良好效果。
综合分析结果表明,在不同土壤条件下,虽然微生物菌肥对番茄青枯病的防治效果存在差异,但施用绿农林 {}^{R}41 号复合微生物菌肥对防治番茄青枯病、改良土壤性状、稳定番茄产量和改善果实品质均有积极的意义。
参考文献
[1]钱程.不同轮作模式对小春作物根际土壤理化性质、微生物和农艺性状的影响[D].昆明:云南农业大学,2023.
[2]郭翠莲.微生物菌肥对油橄榄生长及根际土壤微生物多样性的影响[D].昆明:云南农业大学,2023.
[3]张绪美,曹亚茹,沈文忠,等.微生物肥对设施土壤次生盐渍化和番茄生产的影响[J].中国土壤与肥料,2019(5):119-126.
[4] 岳明灿,王志国,陈秋实,等.减施化肥配施微生物菌剂对番茄产质量和土壤肥力的影响[J.土壤,2020,52(1):68-73.
[5]周开胜.强还原土壤消毒防控连作障碍效果及影响因素研究[D].南京:南京师范大学,2021.
[6] 王杰,龙世芳,王正文,等.番茄青枯病防治研究进展[.中国蔬菜,2020(1):22-30.
[7] 毛骁,孙保平,张建锋,等.微生物菌肥对干旱矿区土壤的改良效果[J].水土保持学报,2019,33(2):201-206.
[8]刘艳霞,李想,曹毅,等.抑制烟草青枯病型生物有机肥的田间防效研究[J.植物营养与肥料学报,2014,20(5):1 203-1 211.
[9]吴晓宗,王岩.生物有机肥防治烟草青枯病及对土壤微生物多样性的影响[J.中国土壤与肥料,2019(4):193-199.
[10]张敏硕,赵英男,杨威,等.微生物菌剂对张北冷凉坝上地区马铃薯产量、品质及活化土壤磷钾的效果[川.水土保持学报,2019,33(3):235-239.
[11] 朱旭.微生物菌剂(水剂)在番茄上的肥效试验[].新农业,2023(15): 16-17.
[12]穆贞,胡雪峰,程畅,等.白叶发酵肥施用对土壤肥力和番茄生长的影响[].土壤通报,2016,47(5):1203-1210.
[13]王晓婷.不同微生物菌肥处理对黄瓜生长及细菌性茎软腐病发生的影响[D].泰安:山东农业大学,2022.
[14]杨威,赵英男,张敏硕,等.温室番茄施用菌剂的促生防病效应[J].河北农业大学学报,2022,45(4):57-63.
[15]潘江禹.枯草芽胞杆菌 BSA-6 防控香蕉枯菱病初步研究[D].海口:海南大学,2013.
[16]张霞.“复合微生物肥料”在小白菜上的肥效试验[].河南农业,2021(1):19.
[17]于建,宋以玲,丁方军,等.复合微生物肥料对盐胁迫下番茄生理特性的影响[].土壤通报,2020,51(2):372-380.
[18]张娜,黄炎,徐谓,等.移栽定殖根际有益菌番茄苗的田间效应研究[J].土壤,2019,51(4):658-664.
[19]赵玲玉,索升州,赵祺,等.梭梭根际促生菌(PGPR)菌肥对番茄产量、品质和土壤特性的影响[.甘肃农业大学学报,2022,57(3): 42{-}51{+}57
[20]李静.土壤有机质测定方法比对分析[].绿色科技,2012,14(5): 203-204.
[21]吴才武,夏建新,段峥嵘.土壤有机质测定方法述评与展望[J].土壤,2015,47(3):453-460.
[22]熊明华,曹焰晖,杨芮,等.一种改良的可直接用于PCR扩增的土壤 DNA 提取方法[].基因组学与应用生物学,2019, 38(4): 1 643-1 648.
[23]曾莉莎,王芳,周海琪,等.大蕉枯菱病菌二重PCR 分子快速检测技术的建立[].南方农业学报,2023,54(8):2 352-2 359.
[24]杜文英.烟草青枯雷尔氏菌和疫霉菌分子检测及土壤微生物分析[D].昆明:云南农业大学,2023.
[25]何子康,张纪利,聂锦瑶,等.土壤中青枯病菌快速检测方法的建立与应用[].中国植保导刊,2022,42(8):5-9+14.
[26]姜伊琳.人参锈腐病菌 Ilyonectria robusta qPCR 检测方法的建立及应用研究[D].长春:吉林农业大学,2023.
[27]王晖,郭云峰.几种土壤速效养分快速测定方法的比较研究[J].天津农林科技,2000(2):9-11.
[28]李得铭,翟子翔,邓涛,等.番茄青枯菌分离与三重PCR体系建立[J.分子植物育种,2020,18(11):3655-3 661.
[29]刘红明,王绍华,郑玉龙,等.3种砧木嫁接对柠檬幼树光合特性的影响[]].经济林研究,2018,36(1):29-34.
[30]胡发广,刘红明,毕晓菲,等.云南干热河谷区7个小粒咖啡叶片光合特性日变化的研究[J.江西农业学报,2020,32(10): 52-56.
[31]何嘉伟.基于叶绿素a测定的监测方法研究与改进[].工业微生物,2023,53(6):1-3.
[32]周春水,莫淋琳,岑江霞,等.不同遮荫条件和栽培基质对蜘蛛抱蛋属植物光合作用的影响[J].北方园艺,2024(7):50-56.
[33]董建舒,申孝军,衣若晨,等.花生叶面积指数精准快速监测方法研究[J].节水灌溉,2024(2): 88{-}94{+}104
[34]马荣辉,梁金英,王宪刚,等.化肥减量配施有机肥对设施番茄产量、品质和土壤肥力的影响[.北方园艺,2023(24): 45-51.
[35]陈琪,董静,周伟伟,等.腐植酸钾和氨基酸肥料对砂培番茄生长与果实品质的影响[J].华北农学报,2023,38(S1):300-306.
[36]刘中良,高俊杰,陈震,等.氮素形态配比对设施基质栽培番茄产量及品质的影响[J].中国蔬菜,2023(7):63-67.
[37]谭洪吉,高艳明,李建设,等.不同功能肥料对基质栽培樱桃番茄的品质产量及基质环境的影响[].浙江大学学报(农业与生命科学版),2022,48(4):434-442.
[38]王晓龙,杨嬰,李红,等.低温对7个首品种种子萌发的影响[J].种子,2022,41(5): 93-97.
[39]邵雪凤.防控土传病害的绿农林 ((\omega)41 号菌肥田间激活技术研究与应用[D].武汉:华中农业大学,2020.
[40]郭立佳,汪军,杨腊英,等.含氨基酸水溶肥料对2株芽孢杆菌的激活作用[].热带作物学报,2020,41(3):549-555.
[41]张洁梅,张仁军,姚正平,等.烟草青枯病生防菌的筛选及其田间防效评价[].中国农学通报,2020,36(28):131-136.
[42]张国庆.生防菌对水稻幼苗生长和根际土壤酶影响的研究[D].大庆:黑龙江八一农垦大学,2012.
[43]宛甜甜.解淀粉芽孢杆菌 SN16-1对水稻稻瘟病的防治作用及其诱抗机理研究[D].上海:华东理工大学,2023.
[44]柳慧静.两种微生物菌种处理对盆栽辣椒的促生作用研究[].南方农业,2022,16(3): 48{-}50{+}56
[45]李舒展.生防菌的鉴定及其对黄瓜枯菱病的防治和对生长的促进作用[D].南京:南京农业大学,2017.
[46]汪军,周游,杨腊英,等.施用复合菌肥与套作对香蕉枯萎病控病作用的影响[J].中国果树,2019(6):69-72+86.
[47]罗静静,刘小龙,李克梅,等.几种微生物菌剂对连作棉田枯黄萎病的防病效应[].西北农业学报,2015,24(7):136-143.
[48]王丽丽,朱诗君,狄蕊,等.微生物菌肥菌剂对番茄生长发育和产量品质的影响[].土壤与作物,2022,11(1):88-95.
黎药胆木根际土壤真菌群落结构分析
霍天祺 罗崇一 杨瑞菊 袁彪 徐颖瑞杨卫丽(海南医科大学海南海口571199)
摘要植株根际土壤微生态群落与植物的生长发育及代谢密切相关,真菌作为微生物区系重要的组成部分,对植株的生长发育也至关重要。本研究提取胆木植株根际土壤真菌菌群总DNA,PCR后使用高通量测序平台对其进行ITS序列测序。根据有效数据统计、ASV分布统计及Alpha分析进行样品验证;根据样品真菌菌群结构分析可得知,胆木根际土壤真菌群落主要聚集在子囊菌门的漆斑霉属,其次在担子菌门、球囊菌门等菌门也丰度较高。微生物对植物的影响一直是研究热点,胆木根际土壤真菌微生态群落结构分析,可为真菌类微生物与胆木植株间的相关性研究提供一定参考。
关键词胆木;根际土壤真菌;菌群结构
中图分类号S533 文献标识码A DOI:10.12008/j.issn.1009-2196.2025.02.009
Analysis of the Rhizosphere Soil Fungal Community Structure of LiyaoNauclea officinalis
HUO Tianqi LUO Chongyi YANG Ruiju YUAN Biao XU Yingrui YANG Weili (Hainan Medical University, Haikou, Hainan 571199, China)
Abstract The rhizosphere soil microecological community is closely related to the growth and metabolism of plants.Fungi, as a vital part of the microbial community, are also crucial to the growth and development of plants. In this study, the total DNA of the fungal community in the rhizosphere soil of the Nauclea officinalis was extracted, and the ITS sequence was sequenced via a high-throughput sequencing platform after PCR. According to the effective data statistics, including ASV distribution statistics and alpha analysis for sample verification, and according to the analysis of the fungal community structure of the sample, it can be known that the fungal community in the rhizosphere soil of N_{\ast} officinaliswasmainlyconcentratedin the Myrothecium of Ascomycota,followed by Basidiomycota, Glomeromycota and other phyla.The effects of microorganisms on plants have always been a research hotspot. The analysis of its fungal microecological community structure in the rhizosphere soil of N_{*} officinalis can provide some reference for the correlation between fungal microorganisms and N_{*} officinalisplants.
uclea officinalis; fungi in the rhizosphere soil; microbial community stru
胆木为茜草科植物乌檀(NaucleaoficinalisPierre.exPitard)的枝、树皮。胆木主要分布于海南和两广地区,又名山熊胆、熊胆树等,常用于治疗扁桃体炎、支气管炎、咽喉炎等,具有清热解毒、消肿止痛的功效。目前胆木的研究主要集中在化学成分、药理作用、栽培技术等方面。胆木中化学成分类型较多,主要含有生物碱类、类及其皂苷、有机(酚)酸类及其苷、环烯醚类、黄酮类5大类成分-2],以吲哚类生物碱异长春花苷内酰胺为代表的生物碱类成分被认为是其主要活性成分。胆木的药理作用主要集中在抗炎、镇痛、抗菌、抗病毒、抗肿瘤、杀疤原虫等3-5]。目前,市场上有关胆木的制剂也已得到广泛使用,例如胆木浸膏糖浆、胆木注射液等。中药材规范化生产的难点之一是药材质量良不齐,而药材的质量除与基因相关外,还与药用植物的生长环境密切相关,包括温度、湿度、光照、以及药材内生微生态群落等。胆木的发展过程中,人们常常忽略了土壤微生态群落,药材产地,植物内生真菌等因素对胆木药用价值的影响。
近年来,研究者逐步发现,植物内生菌通过与宿主植物间特定的互作关系、构建根际微生物区系以及植物根部分泌的代谢物来促使植物生长和发生防御反应"。根际土壤是指与植物根系紧密联系的土壤区域,植物根际是植物、土壤以及微生物相互作用的重要场所"。植物一土壤一根际微生物三者之间在长期相互影响下形成一个复杂的有机整体。植物的生长状态发生变化,植物周围土壤以及根际微生物群落也会随之发生变化,根际微生物发生改变后改善土壤质量,对促进植物生长发育以及促进植物产生次生代谢产物具有重要意义"。根际微生物作为根际的重要组成部分,在植物与土壤之间的互相关系中以直接或间接的方式起到重要作用。植物根系以及根际微生物都需要通过土壤获得营养物质,根际微生物主要通过植物根系分泌物获得能量,同时根际微生物促进有机质的分解、养分的转化等,帮助植物更好地利用土壤中的营养物质。因此,植物根际微生物也是对植物生长发育产生重要影响的因素之一[10-]。真菌作为微生物区系的重要部分,参与有机质的分解,能量转换等过程,对调节土壤可持续生产力具有重要作用[2]。利用土壤微生物来改善土壤质量以及植物生长发育成为研究热点,通过研究胆木根际中有益微生物群落,对根际土壤质量进行改良,进一步提高胆木产生次生代谢产物的能力,对于胆木药材质量的控制具有深远意义。
本实验以胆木根际土壤为研究对象,运用高通量测序技术对根际土壤微生物群落进行测序,使用生物信息学技术对测序结果进行分析,得到胆木根际土壤真菌群落结构,为探究胆木根际土壤真菌对于宿主植物的影响提供研究基础。
1 材料与方法
1.1 材料
1.1.1主要试剂无菌水, 75% 乙醇, 95% 乙醇,3% 次氯酸钠,PDA,培养基,E.Z.N.A superscript{registered} SoilDNAKit DNA 提取试剂盒,AxyPrepPCR CleanupKit回收试剂盒,琼脂糖凝胶, 1%TE 缓冲液,液氮。1.1.2主要仪器高压蒸汽灭菌锅(南京贝登医疗股份有限公司,L5-75LD),生化培养箱(上海博讯实业有限公司医疗设备厂,BSP-400),冷冻离心机(MicrofugeR22RCentrifuge),涡旋振荡仪(华利达,WH-861VortexShaker),恒温水浴锅(上海博讯实业有限公司,DK-8D),PCR仪(德国艾本德公司,NexusX2),电泳仪(北京六一生物科技有限公司,DYY-8C),凝胶成像仪(上海天能生命科学有限公司,Tanon1600)。
1.2方法
1.2.1样品采集及处理在中国热带农业科学院海口院区( 110°20^{\prime}E 19°59^{\prime}N )采集胆木根际土壤,采集时间为2023年9月。选择6棵生长状态良好的胆木,去除胆木根际地表 4~5~cm 的土壤,取附着在根际 0~5 \mm 的土壤,剔除碎石和植物残根等杂物。采集后迅速用保鲜袋封口,做好相关采集标记置于低温保鲜盒中送回实验室。
1.2.2胆木根际土壤微生物总DNA提取、PCR及纯化将采集得到的根际土壤放人液氮中冷冻3 ~h~ ,使用E.Z.N.A. ^registered SoilDNAKitDNA提取试剂盒提取根际土壤中DNA。对提取得到的总DNA进行ITS区PCR扩增,PCR引物为F(5'-GAAC-CWGCGGARGGATCA-3')/R(5'-GCTGCGTTCTTCATCGATGC-3);PCR体系为 25~\upmuL : Phusion Hotstart flex 2X Master Mix 12.5~\upmuL ,上下游引物各2.5~\upmuL ,模板DNA 50~ng , ddH_{2}O 加至 25~\upmuL 。PCR条件为 98 °C 预变性 30~s 98 °C 变性 10~s , 54 °C 退火 30~s , 72 °C 延伸45s进行32循环; 72 °C 最终延伸 10 \min 。PCR结束后,取 5~\upmuL PCR产物进行2% 琼脂糖凝胶电泳检测,并使用回收试剂盒对目标片段进行回收。
1.2.3上机测序及生物信息学分析回收得到的PCR产物使用Quant-iTPicoGreends DNA AssayKit在Qbit荧光定量系统上对文库进行定量,将合格的上机测序文库进行梯度稀释、经NaOH变性为单链,随后使用NovaSeq测序仪进行 2x 250bp的双端测序。
根据双端序列的重叠区,采用Pear将R1、R2序列拼接成长的tag 序列,并使用cutadapter去除barcode以及引物序列。然后Fqtrim过滤低质量序列,采用Vsearch过滤嵌合体。使用DADA2进行降噪后,得到特征表和特征序列。通过QIIME2流程分析得到Alpha多样性信息,并且与SILVA和unite数据库进行序列比对后得到胆木根际土壤真菌物种注释信息。
2 结果与分析
2.1有效数据统计
通过高通量测序平台对胆木根际土壤真菌总DNA进行测序,得到未经处理的测序结果,即原始数据,有效数据是指对原始数据进行质量控制后得到的数据,有效数据的统计结果见表1。样本数据 Q30% 均大于 85% ,说明6组数据均符合实验要求,可以进行后续分析。
| 样本 | 原始数据 | 有效数据 | 有效率/% | Q20/% | Q30/% | GC/% |
| R_1 | 87259 | 81 494 | 93.39 | 99.04 | 97.17 | 56.52 |
| R_2 | 83912 | 78490 | 93.54 | 99.47 | 98.28 | 57.01 |
| R_3 | 81 161 | 75800 | 93.39 | 99.41 | 98.11 | 55.06 |
| R_4 | 81 601 | 77792 | 95.33 | 99.22 | 97.55 | 54.61 |
| R_5 | 81 749 | 76 246 | 93.27 | 99.16 | 97.79 | 55.54 |
| R_6 | 86 977 | 82735 | 95.12 | 99.44 | 98.11 | 56.00 |
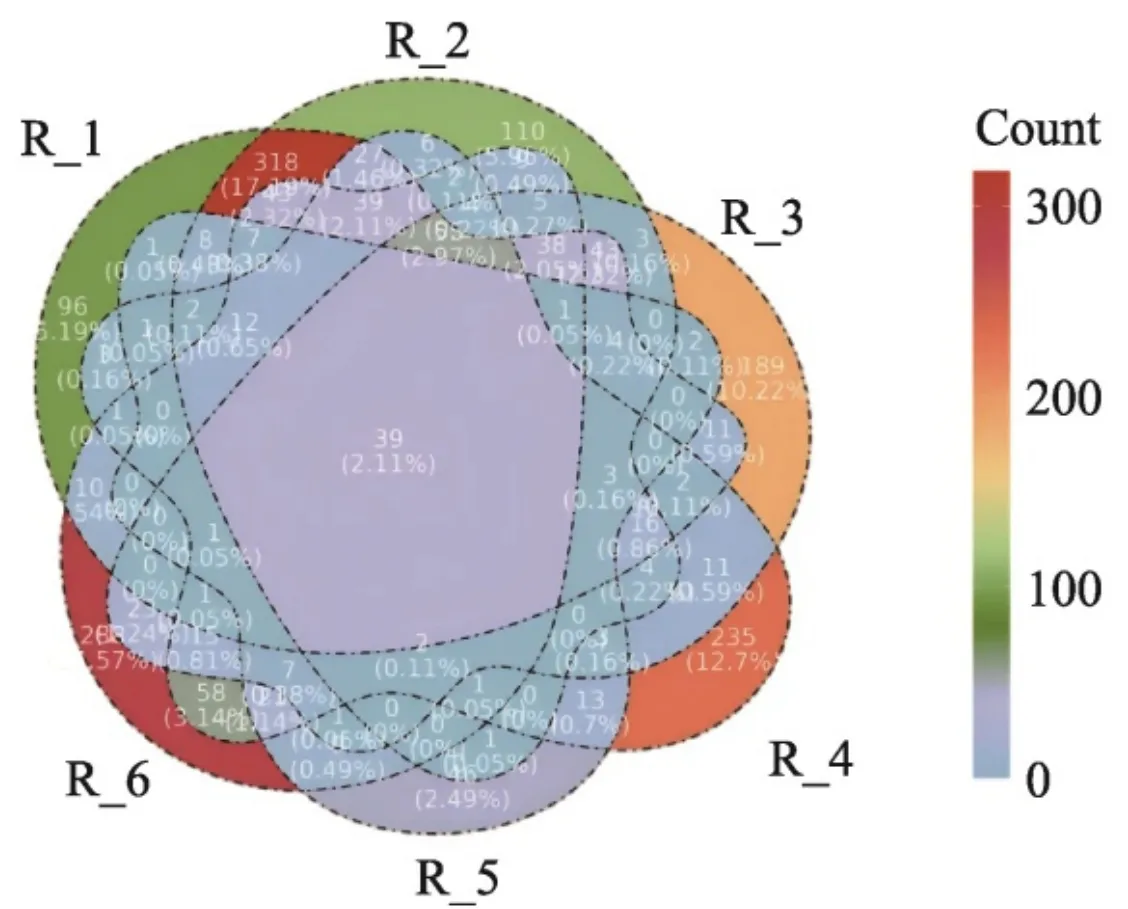
2.2ASV分布Venn图
ASV是测序结果经过去除背景噪音获得的特征序列,对每个样本的ASV进行统计,使用Venn图对统计结果进行展示。由Venn图(图1)可以看出,6个样本的共有特征序列有39个,6个样本各自特有的特征序列数分别为96、110、189、235、46以及288。结果表明,6个样本各自特有的特征序列数大于共有的特征序列数,样本之间有一定的差异性。
2.3Alpha多样性分析
Alpha多样性是指一个特定环境或生态系统中的多样性,常用 Alpha 多样性反映物种丰富度和均匀度以及测序深度。对样品进行 Alpha 多样性分析,由图2可知,R_1,R_2,R_3,R_4,R_6 四组样本表达相较于R_5相对集中,真菌表达密度主要集中在 2.5~5.0 ,6组当中又以R_5的中位线最长,R_1,R_2次之。

2.4胆木根际土壤真菌菌群结构分析
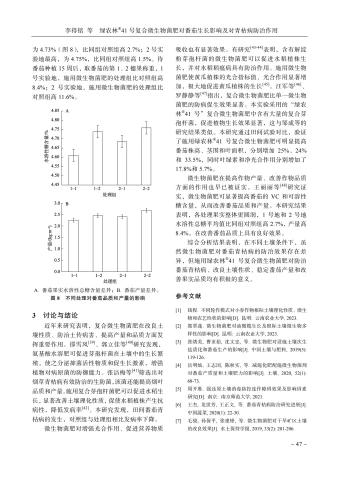
基于物种注释表和物种丰度表,使用柱状堆叠图及气泡图(图3)展示胆木根际土壤真菌丰度TOP30的真菌。由图3-A可知,6组中优势菌门均为Ascomycota,占比在 57.78%(R_{-}4)~79.72% (R_2)。其次主要分布在 Basidiomycota,以R_4占比最高,达到 27.02% , ~R~\_3(12.40%) rm{R} 5(13.24%) 次之。剩余少部分集中于Fungi-unclassified,Glomeromycota 和 Chytridiomycota。R_3 组 Zyg-omycota含量也较高,达到 6.53% 。由图3-B可知,为Myrothecium是 ~R~\_{1}\dots~R~\_{2}\dots~R~\_{3} 组的优势菌属,占比依次为 12.29% 、 13.04% 、 9.48% ,并且在Fungi-unclassified、Bionectria、Humicola、Ascom-ycota-unclassified等菌属丰度也较高,但比例均不同。除 Fungi-unclassified 外,R_4 组优势菌属为Agaricomycetes_unclassified,占比为 17.76% ,其次为Humicola,占比为 11.46% ,除Fungi-unclassified外,前3组的优势菌属Myrothecium在R_4也有较高丰度 (4.88%) 。R_5组优势菌属为Bionectria,占比达到 18.57% ,其次Humicola丰度也较高(13.99%) 。R_6组优势菌属为Fungi-unclassified(12.29%) ,其次为Myrothecium (7.64%) ,Ascom-ycota_unclassified (7.27%) 。由图3-C可得,优势菌属Myrothecium、Bionectria、Humicola、Ascomycota-unclassified在各组中所占比例优势明显,且这些菌属同属Ascomycota菌门。因此,优势菌门Ascomycota在各组均优势明显,占比均达到 50% 以上。其次Basidiomycota菌门中Agaricomycetes_unclassified在R_4组中丰度优势也较为明显。
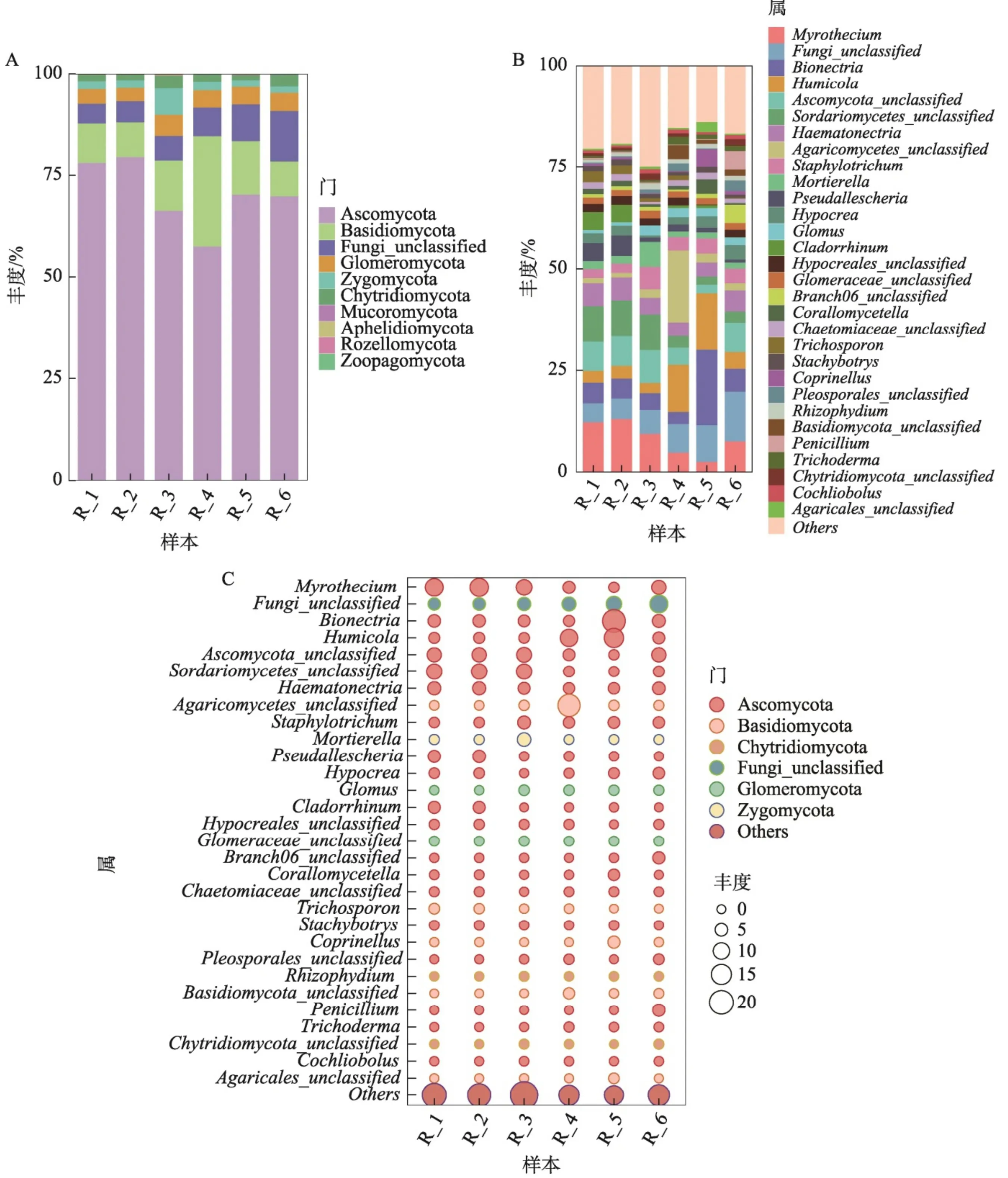
图3胆木根际土壤真菌物种组成分析
3 讨论与结论
高通量测序可规避传统分离培养的盲目性与随机性,能快速准确地了解微生物群落组成结构和生态功能[13]。本研究使用高通量测序平台对胆木根际土壤真菌进行测序,通过数据统计,6组胆木根际土壤真菌共有特征ASV序列39个,各自的特征序列数在46\~288,注释10个门,34个纲,83个目,164个科,291个属,428个种。同时明确了子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、球囊菌门(Glomeromycota)、壶菌门(Chytridiomycota)和接合菌门(Zygomycota)为胆木根际土壤中的优势菌门,漆斑菌属(Myrothecium)、腐质霉属(Humicola)、生赤壳属(Bionectria)等为胆木根际土壤真菌中优势菌属。漆斑菌属为R_1、R_2、R_3组的最优势菌属,占比均在 9% 以上。有研究表明,漆斑霉属真菌疣孢漆斑菌(Myrotheciumverrucaria)是生物防治寄生线虫的重要真菌[4]。此外,更有学者从多种植物中分离得到漆斑霉属菌株,比如地锦草、半夏等,并且其次生代谢产物具有较好的抗白色念珠菌活性5-16]。本研究为后续微生物群落的生态功能预测、优势菌属的进一步发展等奠定了基础。
植物微生物组在营养循环和植物生长中起关键作用,它们的形成受到植物、微生物和环境三者之间复杂相互作用的影响,根际微生物具有促进宿主植物生长发育、防卫病原菌等功能。土壤微生物菌群的失调会大幅度增加植物发病的几率[1]。例如研究发现,豆科植物根际微生态系统中常见的共生微生物丛枝菌根真菌(AMF)和根瘤菌不是孤立地发挥作用,它们在与豆科植物长期共生与进化的过程中,形成了AMF一根瘤菌一豆科植物三者共生的关系。同时接种AMF和根瘤菌相较于单独接种更能促进宿主植物的生长发育,其原因是根瘤菌和部分AMF菌株对宿主植物的影响具有协同效应[18-19]。深入分析植物与微生物的相互作用及作用机制,对更好地利用微生物使其对宿主植物产生正向的影响具有重要意义。同时,植物根系一土壤一微生物之间的相互作用对整个土壤环境的稳定和发展也至关重要[20]。
与药用植物共生的真菌会产生一些诸如抗肿瘤抗氧化等活性成分,而由其诱导或非诱导产生的次生代谢产物,对植物药用成分的形成和含量的影响十分巨大。如龙血树中血竭的形成,白木香树中沉香的形成和红豆杉树紫杉醇的生成等[21-22]。药用植物根际土壤真菌长期与药用植物真菌保持紧密联系,不仅根际土壤真菌会影响药用植物的生长发育以及次生代谢产物的产生,真菌自身的次生代谢产物的形成也会受到影响。植物根际土壤真菌的研究,从植物生长发育,药用价值分析等角度来看,可以发现植物根际土壤真菌与药物疗效之间的影响机制尚未探究清晰。胆木根际土壤真菌群落结构分析,为后续研究胆木次生代谢产物与根际土壤真菌群落的相关性提供研究基础,可研究与胆木次生代谢产物的生成与积累有关的菌群,制造可高效应用于提升胆木药材品质从而对胆木相关制剂疗效产生积极影响的菌物制剂,这将对胆木药材进一步应用产生深远意义。
参考文献
[1]戚卫蕊,王德立,冯锦东,等.南药胆木的研究进展[].安徽农业科学,2016,44(16):111-113.
[2]麦世瑛,王怡然,李永辉,等.中药胆木化学成分及其药理活性研究进展[J].广州化工,2018,46(16):38-41.
[3] Wang HY, Wang R X, Zhao Y X, et al. Three new isomericindole alkaloids from Nauclea officinalis[J]. Chemistry &Biodiversity, 2015, 12(8): 1 256-1 262.
[4]刘腾,常艳璐,王斌.胆木浸膏糖浆的药理作用与临床应用研究进展[J].药品评价,2020,17(16):14-17.
[5]Sun JY, Lou H X, Dai S J, et al. Indole alkoloids fromNauclea ofcinalis with weak antimalarial activity[J].Phytochemistry, 2008, 69(6): 1 405-1 410.
[6] 王月,廖锦红,孙立新.胆木及其制剂研究进展[川].亚太传统医药,2018,14(8):80-84.
[7]Hu L F, Robert C A M, Cadot S,et al. Root exudate metabo-lites drive plant-soil feedbacks on growth and defense byshaping the rhizosphere microbiota[J]. Nature Communica-tions, 2018, 9(1): 2 738.
[8]阿拉坦存布尔,曹乌吉斯古楞,包金花,等.手参内生真菌及根际土壤真菌多样性研究进展[川.现代农业科技,2023(21): 76-83+86.
[9]Philippot L, Raaijmakers J M, Lemanceau P, et al. Goingback to the roots: the microbial ecology of the rhizosphere P[J]. Nature reviews. Microbiology, 2013, 11(11): 789-799.
[10]Qu Q,Zhang Z Y Y,Peijnenburg W J G M, et al.Rhizosphere microbiome assembly and its impact on plantgrowth[J]. Journal Of Agricultural And Food Chemistry,2020, 68(18): 5 024-5 038.
[11]Finkel O M, Castrillo G,Herrera Paredes S,et al.Under-standing and exploiting plant beneficial microbes[J]. CurrentOpinion in Plant Biology, 2017, 38: 155-163.
[12]王晓春,高婷.盐碱地紫花首根际土壤真菌多样性分析[J].农业与技术,2023,43(24):9-12.
[13] Mishra S, Goyal D,Phurailatpam L.Targeted 16S rRNAgene and ITS2 amplicon sequencing of leaf and spike tissuesof Piper longum identifies new candidates for bioprospectingof bioactive compounds[J]. Archives of Microbiology. 2021,203(7): 3 851-3 867.
[14]董海龙,王海香,张作刚,等.抑杀根结线虫的疣孢漆斑菌代谢活性物质的分离鉴定.植物保护学报,2019,46(3): 721-722.
[15]姜松杰.两株大戟属植物内生真菌的次级代谢产物及抗菌活性研究[D].兰州:兰州大学,2024.
[16]虞菊萍,杨鸣华,何小荣.一株植物内生真菌漆斑菌次级代谢产物的研究[J].中国药科大学学报,2021,52(4):431-437.
[17]寇智瑞,周鑫斌.不同连作年限黄壤烟田土壤细菌群落的差异[J].植物营养与肥料学报,2020,26(3):511-521.
[18]段海霞,师茜,康生萍,等.丛枝菌根真菌和根瘤菌与植物共生研究进展[J/OL].草业学报:1-17[2024-01-31].http://kns.cnki.net/kcms/detail/62.1105.S.20240108.0850.014.html.
[19]刘忆,袁玲.根瘤菌和AM真菌对紫花首结瘤和产质量的影响[J].土壤学报,2020,57(5):1292-1298.
[20]连文慧,董雷,李文均.土壤环境下的根际微生物和植物互作关系研究进展[J].微生物学杂志,2021,41(4):74-83.
[21]Stierle A,Strobel G, Stierle D. Taxol and taxane productionby Taxomyces andreanae, an endophytic fungus of Pacificyew[J]. Science, 1993, 260(5 105): 214-216.
[22]杨涛,赵疆,魏亚琴,等.华重楼内生真菌聚多曲霉的促生与拮抗作用[J].微生物学通报,2021,48(8):2665-2680.
两种PCR快速检测转基因糜子中外源基因的插人拷贝数
董晓静1.2 夏启玉'降彦苗”刘国庆”程汝宏’赵辉1(1.中国热带农业科学院三亚研究院/中国热带农业科学院热带生物技术研究所/海南省南繁生物安全与分子育种重点实验室海南三亚572024;2.洛阳师范学院生命科学学院河南洛阳471934;3.河北农林科学院谷子研究所河北石家庄050035)
摘要近年来,糜子(PanicummiliaceumL.)的转基因育种研究越来越多,本研究旨在建立简单快速检测转基因糜子中外源基因插人拷贝数的方法。从糜子基因组中筛选了一个单拷贝基因SDI,并以其为内参基因,以潮霉素抗性基因(Hyg)为外源基因,建立了数字PCR检测转基因糜子中外源基因插人拷贝数的方法。并以数字PCR鉴定的一个双拷贝样品为参照样本,建立了荧光定量PCR检测转基因糜子中外源基因插入拷贝数的方法。生物信息学分析表明,SDI为单拷贝基因,可作为转基因糜子中外源基因插入拷贝数检测的内参基因。数字PCR法检测的转基因糜子样品中,9份样品均为单拷贝或双拷贝插人;荧光定量PCR法检测的转基因糜子样品中,10份样品为单拷贝或双拷贝插人,其余13份样品均为多拷贝插人;对其中的4份低拷贝样品也进行了数字PCR检测,结果与荧光定量PCR法的检测结果完全一致,表明荧光定量PCR法检测转基因糜子中外源基因低拷贝时的准确度较高。本研究建立的数字PCR法简单快速,准确度高,适合在样品数量较少时使用,而荧光定量PCR法检测低拷贝插人时准确度较高,且成本低廉,适合从大量转基因株系中初筛出的低拷贝插人株系。这2种方法均可简单快速地检测糜子转基因育种中外源基因插人拷贝数。
关键词转基因糜子;外源基因;插人拷贝数;数字PCR;荧光定量PCR 中图分类号S516 文献标识码 A DOI: 10.12008/j.issn.1009-2196.2025.02.010
Rapid Detection of the Insertion Copy Number of Exogenous Genes in Transgenic Broomcorn Millet via Two PCR Methods
DONG Xiaojing12 XIA Qiyu' XIANG Yanmiao? LIU Guoqing? CHENG Ruhong? ZHAO Hui' (1. Sanya Research Institute, Chinese Academy of Tropical Agricultural Sciences/Tropical Bioscience and Biotechnology, Chinese Academy of Tropical Agricultural Sciences/Hainan Key LaboratoryforBiosafety Monitoring and Molecular Breeding in Off-Season Reproduction Regions, Sanya, Hainan 572024, China; 2. College of Life Science, Luoyang Normal University,Luoyang,Henan 471934, China; 3. Institute of Millet Crops,Hebei Academy of Agriculture and Forestry Sciences,Shijiazhuang,Hebei 050035,China)
Abstract In recent years, there has been an increasing amount of research on the transgenic breeding of broomcorn millet (Panicum miliaceum L.). This study aimed to establish simple and rapid methods for detecting the insertion copy number of exogenous genes in transgenic broomcorn millet. This study screened the single-copy gene SDl from the genome of broomcorn millet and established a digital PCR method to detect the insertion copy number of exogenous genes in transgenic broomcorn millet, using it as an internal reference gene and the hygromycin resistance gene (H y g) asanexogenousgene.A fluorescence quantitative PCR method was established to detect the insertion copy number of exogenous genes in transgenic broomcorn millet, using a double copy sample identified by digital PCR as the reference sample. Bioinformatics analysis shows that SDl is a single copy gene and can be used as an internal reference gene for insertion copy number detection of exogenous genes in transgenic broomcorn millet; Among the transgenic broomcorn millet samples detected by digital PCR, nine samples were either single copy or double copy insertions; Among the transgenic broomcorn millet samples detected by fluorescence quantitative PCR method, ten samples were single copy or double copy insertions, while the remaining 13 samples were multi-copy insertions.Among them,four low-copy number samples were also detected by digital PCR,and the detection results were completely consistent with those of the quantitative fluorescence PCR method,indicating that the quantitative fluorescence PCR method has high accuracy in detecting low-copy number insertions of exogenous genes in transgenic broomcorn millet.The digital PCR method established in this study is simple,fast,highly accurate, and suitable for use when the sample quantity is relatively small. On the other hand, the quantitative fluorescence PCR method has high accuracy in detecting low-copy insertions, and its cost is low, making it suitable for screening low-copy insertions from many transgenic strains. Both methods can detect simply and rapids methods for detecting the insertion copy number of exogenous genes in the transgenicbreedingofbroomcornmillet.
Keywords transgenic broomcorn millet; exogenous genes; insertion copy number; digital PCR; quantitative fluorescence PCR
糜子(PanicummiliaceumL.),又名黍稷,属禾本科黍属,起源于我国的黄河流域,是人类最早驯化的谷类作物之一,具有生育周期短、耐干旱和耐贫瘠等特点,其种植历史已超过了10000年,在俄罗斯、印度、中东和欧洲先后被广泛种植过,目前糜子的种植地区主要在欧洲和亚洲的半干旱地区[-5]。糜子的籽粒富含蛋白质、淀粉、维生素、必需氨基酸、不饱和脂肪酸、微量元素、膳食纤维和酚类物质,还具有预防心脑血管疾病和调节血糖的作用,具有较高的营养价值。糜子是重要的南繁作物,作为一种C4植物,其蒸腾速率低,水分利用率高,能在干旱、盐碱等较差环境条件中生存,且产量相对稳定。因此,糜子常作为备荒作物种植于盐渍化和干旱等自然条件较差的地区[7-8]
在1979年之前,我国的每个省几乎都有糜子栽培,包括海南琼海。糜子曾经是我国北方比较重要的谷类栽培作物,但由于糜子的产量增加缓慢,品质提升难度较大,逐渐变成了地区特色作物"。与玉米、水稻等作物相比,糜子的驯化水平低,还具有许多野生性状,这些野生性状严重阻碍了糜子的育种进程和品种改良。目前我国糜子的育种方法主要为杂交育种和诱变育种,均为常规育种方法,利用现代分子标记辅助育种的研究较少[9-10]。现代基因工程技术的快速发展,为糜子的品种改良提供了快速有效的途径。随着糜子遗传转化技术的成熟,越来越多的学者开始以性状改良为目的进行转基因糜子生物育种方面的研究。2024年,Liu等[敲除了糜子的八氢番茄红素脱氢酶(PDS)基因,得到的转基因糜子植株出现白化症状。同年,Lu等[2也敲除了糜子品种Longmi4的PmSD1基因,获得了植株明显矮化的转基因糜子。
在当前的转基因植物育种研究中,通常会选择低拷贝外源基因插入整合到受体基因组的株系,以用于后续育种研究,这是因为外源基因能否在受体植物中高效表达及稳定遗传与其以低拷贝还是多拷贝插人到受体植物基因组中有关[13]。当插人整合到受体植物的外源基因组是低拷贝(1\~2个)时,通常能高效表达,并且能稳定遗传[14-15]。然而,要想从大量 T_{0} 代转基因株系中筛选出低拷贝插入的转基因株系,需要花费大量的时间。因此,简单快速且容易批量操作的外源基因插人拷贝数检测方法对转基因植物育种研究具有举足轻重的作用。
在检测外源基因插人到受体植物基因组中拷贝数的方法中,最准确的是Southern杂交法,但该方法成本高,周期长,很难实现简单快速且大批量检测转基因植物中外源基因的插人拷贝数。为了建立可从大量转基因糜子株系中快速筛选出低拷贝插人的转基因糜子株系的方法,以便用于后续育种研究,本研究利用生物信息学分析从糜子基因组中筛选出了单拷贝基因SDI,以其作为拷贝数检测的内参基因,利用数字PCR(digitalPCR,dPCR)鉴定了多个单拷贝和双拷贝株系;并以其中的双拷贝株系作为参照样本,建立了基于SYBR染料法的快速筛选外源基因低拷贝插人的转基因糜子植株的荧光定量PCR(fluorescencequantitativePCR,qPCR)方法,这2种方法均可简单快速地检测出转基因糜子植株中外源基因插人的拷贝数。
1 材料与方法
1.1材料
1.1.1植物材料转基因糜子植株共38株( JS1~ JS38),来自中国热带农业科学院三亚研究院。
1.1.2试剂耗材植物DNA通用试剂盒(D3187-02)购自广州美基生物科技有限公司;PCR平盖八排管购自Labselect公司;qPCRPremix(Universal)购自江苏愚公生命科技有限公司;GreenMasterMix购自北京艾德莱生物科技有限公司;dPCR样本制备通用耗材(微滴生成油、密封垫、可穿刺热封膜、微滴生成芯片、密封剂)和微滴PCR预混液均购自广东永诺医疗科技有限公司。引物和探针均由生工生物工程(上海)股份有限公司合成。
1.2方法
1.2.1SD1基因在糜子基因组中拷贝数的生物信息学分析SD1基因,登录号为PM13G15420,来自糜子chr_13染色体,下载该基因核苷酸序列。使用makeblastLongmi-V2版本的pep文件建库,使用SDI基因序列作为查询序列,利用Blast比较查询序列与目标数据库中的序列,找出二者之间的局部相似性区域,最终获得比对结果。
1.2.2内参基因和外源基因的引物设计和PCR扩增条件以SDI基因作为内参基因,以转基因糜子常用的筛选标记潮霉素抗性基因 _{H y g} 作为外源基因,使用Premier5.0软件设计SD1基因和Hyg基因的特异性引物与探针,序列见表1。以1株转基因阳性糜子植株样品的基因组DNA为模板,分别采用SD1-F/SD1-R和Hyg-F/Hyg-R引物对进行温度梯度PCR反应,退火温度分别是52.7、55.6、57.8、60.0、62.2、64.4、66.6、68.4、69.5和 70.0 °C ,实验重复3次。PCR反应体系为:GreenMaster Mix 12.50~\upmuL ,浓度为 10.0\ \upmumol/L 的SD1-F/Hyg-F 1.0~\upmuL ,浓度为 10.0\ \upmumol/L 的SD1-R/Hyg-R 1.0~\upmuL ,浓度为 50.0~ng/\upmuL 的糜子样品DNA模板 1.0~\upmuL ,无菌蒸馏水 9.50~\upmuL ,总体积 25.0~\upmuL 。PCR扩增条件: 94 °C 预变性 5\;min 94 °C 变性 30~s ,退火 30~s , 72 °C 延伸 30~s ,共35个循环;最后 72 °C 延伸 7~min 。分别取 5.0\ {\upmuL} S D I 和 _{H y g} 基因的PCR产物,在 2.0% 的琼脂糖凝胶上进行电泳检测。若 S D I 和 _{H y g} 的PCR 产物电泳检测结果分别出现111和 143~bp 的条带,则为目的基因的扩增条带。将剩余的PCR产物送擎科生物科技股份有限公司测序,将测序结果与SDI基因和 _{H y g} 基因的核苷酸序列进行Blast比对,验证引物是否特异。
| 基因 | 引物名称 | 引物序列(5'-3') | 大小/bp |
| SD1 | SD1-F | CAAGCTCCCGTGGAAGG | 111 |
| SD1-R | CCCATTGGCTCGAAATCCT | ||
| SD1-P | VIC-AGACCCTCTCCTTCGGCTTCCA-MGB | ||
| Hyg | Hyg-F | GTCAAGACCAATGCGGAGCA | 143 |
| Hyg-R | CCCAATACGAGGTCGCCAAC | ||
| Hyg-P | FAM-TCGAAGTAGCGCGTCTGCTGCTCCA-BHQ1 |
1.2.3转基因糜子阳性植株的筛选采集38株转基因糜子植株(JS1\~JS38)的叶片,提取基因组DNA,通过 Hy g 基因的PCR鉴定排除非转基因的糜子植株。PCR扩增体系及程序同1.2.2,退火温度为 60.0 °C 。PCR扩增反应结束后,取 5.0~\upmuL PCR产物在 2% 琼脂糖凝胶上进行电泳检测,若样品扩增出 143~bp 的目的条带,则为转基因阳性糜子植株。
1.2.4dPCR检测转基因糜子中外源基因插入拷贝数从糜子转基因阳性植株中随机选择了9株(JS1、JS3、JS4、JS6、JS25、JS27、JS28、JS29和JS38)作为待测样品。以SDI基因作为内参基因,以 Hy g 基因作为外源基因,引物及探针详见表1。利用dPCR技术检测外源基因在转基因糜子基因组中的插人拷贝数,每个样品进行3个重复,以糜子非转基因植株的基因组DNA为阴性对照。使用MicroDrop微滴式数字PCR系统,反应体系为:dPCR反应预混液 10.0~\upmuL ,浓度为 10.0 \upmumol/L 的内参基因SDI和外源基因 _{H y g} 的正向引物和反向引物各 1.8~\upmuL ,浓度为 10.0\ \upmumol/L 的内参基因SDI和外源基因 _{H y g} 的探针各 0.5~\upmuL ,浓度为50.0\ ng/\upmuL 的糜子样品DNA模板 1~\upmuL ,无菌蒸馏水 0.8~\upmuL ,总体积共 20.0~\upmuL_{o} PCR 扩增程序如下:95 °C 变性 10 \min 95°C~30~s , 60°C\ ~l~ \operatorname*{min} ,进行45个循环; 98 °C\ 10\ min 。PCR扩增反应结束后,将PCR扩增产物放人MicroDrop生物芯片分析仪中,使用QuantDropSoftware读取每个微滴的VIC(Violet invader)和FAM(Fluorescein amidite)通道的荧光信号强度,根据荧光数据,得出阳性微滴和阴性微滴数量,继而得到检测糜子样品基因组DNA中 Hy g 与SDI的拷贝数( \mathsf{c o p y/\upmu L} ),并计算其拷贝数之比,参考谢秀菊等16的方法确定外源 Hy g 基因在糜子基因组DNA中的插人拷贝数。
1.2.5qPCR检测转基因糜子植株外源基因插入拷贝数
1.2.5.1引物验证随机选择7株阳性转基因糜 子样品和1株非转基因糜子样品的基因组DNA 为模板,进行SYBR染料法的qPCR实验,验证 以内参基因SDI和外源基因 Hy g 的引物(表1) 进行qPCR的有效性。qPCR反应在Quant Studio TM1Plus实时荧光定量PCR仪[赛默飞世尔(上 海)仪器有限公司]中进行,反应体系如下:qPCR Premix(Universal) 10.0~\upmuL ,浓度为 10.0\ \upmumol/L 的 S D I 或 Hy g 的正向和反向引物各 0.4~\upmuL ,浓度 为 50.0\ ng/\upmuL 的糜子样品DNA模板 2.0~\upmuL ,Low Rox dye染料 0.2~\upmuL ,无菌蒸馏水 7.0~\upmuL ,总体积 共 20.0 \upmuL 。每个样品设3次重复。 S D I 或 _{H y g} 基因的qPCR扩增程序: 95 °C 预变性 30 ~s~ 95 °C 变性 10~s , 60 °C 退火 30~s ,进行40个循环;融解 曲线, 95°C~15~s , 60°C~60~s 95°C~15~s
1.2.5.2qPCR检测待测转基因糜子样品的外源基因插入拷贝数以 SDI基因为qPCR的内参基因,以 Hy g 基因为qPCR的外源基因,以dPCR鉴定的插入2个拷贝的转基因糜子作为参照样本,通过相对定量qPCR检测23株转基因糜子样品的外源基因插人拷贝数。qPCR使用的引物、反应体系和扩增程序均与引物验证相同,以糜子非转基因植株的基因组DNA为阴性对照,每个糜子样品设3个重复。待转基因糜子样品的qPCR反应结束后,使用QuantStudioTMDesign&AnalysisSoftwarev1.5.2系统对数据进行分析,采用相对定量的 2^{-\Delta\Delta C_{1}} 法计算转基因糜子样品中外源 Hy g 基因相对于参照样本的拷贝数,其中 C t 为循环阈值, \Delta C t_{\#\#\Pi=C}t_{Hyg}-C t_{S D I} , \Delta C t 参照 \scriptstyle:=Ct_{H y g^{-}} Ct_{S D I} \Delta\Delta C t{=}\Delta C t_{\#\#\"{{m}}}{-}\Delta C t 参照。参考谢秀菊等的方法计算外源 _{H y g} 基因在糜子基因组DNA中的插入拷贝数。
2 结果与分析
2.1SD1基因在糜子基因组中的拷贝数分析
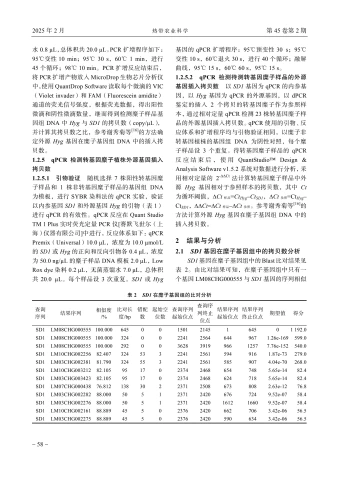
SDI基因在糜子基因组中的Blast比对结果见表2。由比对结果可知,在糜子基因组中只有一个基因LM08CHG000555与SD1基因的序列相似度达 100% ,其余基因与SDI基因的序列相似度最高为 80% 左右。因此,糜子SDI基因为单拷贝基因,可用作转基因糜子中外源基因插人拷贝数检测的内参基因。
| 查询 序列 | 结果序列 | 相似度 /% | 比对长 度/bp | 错配 数 | 起始空 位数 | 查询序列 起始位点 | 查询序 列终止 位点 | 结果序列 起始位点 | 结果序列 终止位点 | 期望值 | 得分 |
| SD1 | LM08CHG000555 | 100.000 | 645 | 0 | 0 | 1501 | 2145 | 1 | 645 | 0 | 1 192.0 |
| SD1 | LM08CHG000555 | 100.000 | 324 | 0 | 0 | 2241 | 2564 | 644 | 967 | 1.26e-169 | 599.0 |
| SD1 | LM08CHG000555 | 100.000 | 292 | 0 | 0 | 3628 | 3919 | 966 | 1257 | 7.76e-152 | 540.0 |
| SD1 | LM10CHG002256 | 82.407 | 324 | 53 | 3 | 2241 | 2561 | 594 | 916 | 1.87e-73 | 279.0 |
| SD1 | LM03CHG002381 | 81.790 | 324 | 55 | 3 | 2241 | 2561 | 585 | 907 | 4.04e-70 | 268.0 |
| SD1 | LM10CHG003212 | 82.105 | 95 | 17 | 0 | 2374 | 2468 | 654 | 748 | 5.65e-14 | 82.4 |
| SD1 | LM03CHG003423 | 82.105 | 95 | 17 | 0 | 2374 | 2468 | 624 | 718 | 5.65e-14 | 82.4 |
| SD1 | LM07CHG000438 | 76.812 | 138 | 30 | 2 | 2371 | 2508 | 673 | 808 | 2.63e-12 | 76.8 |
| SD1 | LM03CHG002282 | 88.000 | 50 | 5 | 1 | 2371 | 2420 | 676 | 724 | 9.52e-07 | 58.4 |
| SD1 | LM03CHG002276 | 88.000 | 50 | 5 | 1 | 2371 | 2420 | 1612 | 1660 | 9.52e-07 | 58.4 |
| SD1 | LM10CHG002161 | 88.889 | 45 | 5 | 0 | 2376 | 2420 | 662 | 706 | 3.42e-06 | 56.5 |
| SD1 | LM03CHG002275 | 88.889 | 45 | 5 | 0 | 2376 | 2420 | 590 | 634 | 3.42e-06 | 56.5 |
2.2SD1和 ±b{H}±b{y}±b{g} 基因引物PCR扩增条件摸索
利用温度梯度PCR摸索内参基因SDI和外源基因 Hy g 的引物扩增的最适退火温度,PCR产物的电泳检测结果如图1所示。退火温度为52.7、55.6、57.8、60.0、62.2和 64.4 °C 时,SDI基因和Hyg 基因均能扩增出目的条带,但在退火温度为60.0 °C 时条带最亮,且无非特异性条带。经测序比对可知,SDI基因和 Hy g 基因的PCR产物分别与糜子SDI基因和外源 _{H y g} 基因的序列完全一致,因此SD1-F/SD1-R引物对可用于糜子SD1基因的检测,Hyg-F/Hyg-R引物对可用于外源基因Hyg基因的检测,均选择退火温度 60.0 °C

M.DNA分子量标准MarkerDL2000,单位为bp,从上往下分子量依次为2000、1000、750、500、250和100,下同。编号1\~10,依次是52.7、55.6、57.8、60.0、62.2、64.4、66.6、68.4、69.5和 70.0 °C
2.3转基因糜子阳性植株的筛选
38株糜子植株的PCR检测结果见图2。其中,JS1、JS3、JS4、JS6-JS10、JS14-JS19、JS21-JS25、JS27-JS34和JS38为转基因阳性糜子植株,共28株;JS2、JS5、JS11-JS13、JS20、JS26、JS35-JS37为非转基因糜子植株,共10株。
2.4dPCR法检测待测样品中外源基因插入拷贝数
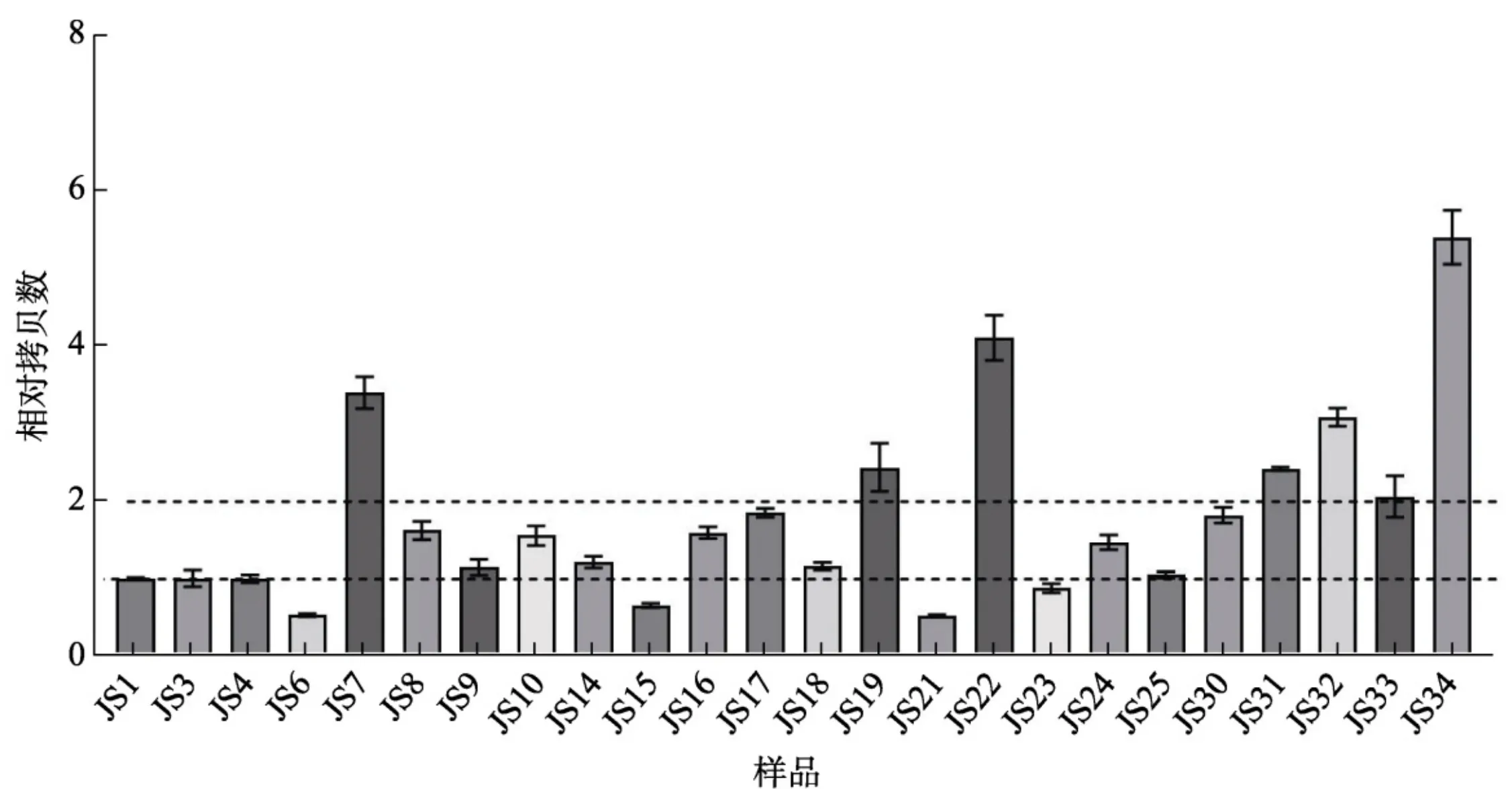
9份转基因糜子样品的dPCR散点图见图3。9份样品的反应中形成的微滴数均大于50000,满足dPCR对于微滴数量的分析要求。图中的红线为阈值,可明确区分 Hy g 基因与SDI基因的阴性微滴和阳性微滴,读出 Hy g 基因与SD1基因的阴性微滴和阳性微滴的数目。根据泊松分布的原理,通过阳性微滴的比例计算出目标核酸分子的绝对拷贝数,再计算出待测糜子样品的Hyg基因与SD1基因的拷贝数比值(表3),根据3次重复实验的拷贝数比值的平均值计算外源基因插入拷贝数。结果显示,JS6的拷贝数比值接近0.5,为单拷贝插人植株;JS1、JS3、JS4、JS25、JS27、JS28、JS29和JS38的拷贝数比值接近1,为双拷贝插人植株。

图2转基因糜子阳性植株的PCR鉴定
2.5qPCR法检测待测样品中外源基因插入拷贝数
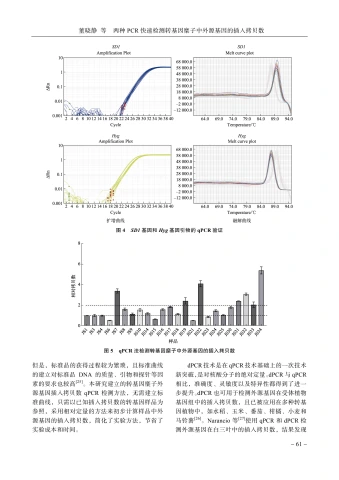
2.5.1引物验证SD1基因和Hyg基因引物的qPCR的扩增曲线及融解曲线(图4)表明,2个基因的引物均有明显的扩增曲线,且融解曲线均具有唯一的吸收峰,说明这2对引物扩增效率良好,且无非特异性扩增。因此,这2对引物可作为SDI基因和Hyg基因的SYBR法qPCR引物。
2.5.2qPCR法检测样品中外源基因插入拷贝数23份转基因糜子样品外源基因插人拷贝数的qPCR检测结果见图5。其中,3份样品(JS6、JS15和JS21)的Hyg基因的相对拷贝数约为0.5,是单拷贝插人的转基因植株;7份样品(JS3、JS4、JS9、JS14、JS18、JS23和JS25)的 _{H y g} 基因的相对拷贝数约为1,是双拷贝插人的转基因植株;其余样品均为三拷贝及以上插人的转基因植株。其中,对单拷贝样品(JS6)和双拷贝样品(JS3、

图3SD1基因和 _{H y g} 基因的dPCR散点图
| 样品 | Hyg拷贝数/SDI拷贝数比值 | 外源基因插 人拷贝数 | |||
| 重复1 | 重复2 | 重复3 | 平均值 | ||
| JS1 | 0.91 | 0.88 | 0.85 | 0.88 | 2 |
| JS3 | 1.07 | 1.14 | 0.97 | 1.06 | 2 |
| JS4 | 0.91 | 1.13 | 0.88 | 0.97 | 2 |
| JS6 | 0.40 | 0.40 | 0.42 | 0.41 | 1 |
| JS25 | 0.81 | 0.84 | 0.77 | 0.81 | 2 |
| JS27 | 0.95 | 0.77 | 0.80 | 0.84 | 2 |
| JS28 | 1.09 | 0.93 | 1.15 | 1.06 | 2 |
| JS29 | 0.96 | 0.96 | 0.90 | 0.94 | 2 |
| JS38 | 0.84 | 0.82 | 0.82 | 0.83 | 2 |
JS4和JS25)也进行了dPCR 检测,检测结果与qPCR检测的结果完全一致,说明qPCR法检测外源基因单、双拷贝插人时的准确度较高。
3 讨论与结论
3.1讨论
qPCR技术作为一种DNA定量检测的经典方法,被广泛应用于检测转基因植株中外源基因插人到受体基因组的拷贝数。目前,qPCR已经成功应用于转基因水稻[7-19]、棉花[20]、马铃薯[2]、柑橘[22]、番茄[23]和烟草[24]等作物的拷贝数检测。例如,魏毅东等[9]利用基于 Taqman 的多重qPCR成功检测了10株转基因水稻材料的外源基因插入拷贝数。任爽等[23]利用基于 Taqman 的qPCR成功检测了12株PDS基因编辑番茄材料的外源基因插人拷贝数。铁原毓等[24]利用基于SYBRGreen的qPCR成功检测了14株转基因烟草材料的外源基因插人拷贝数。以上方法需要利用已知转基因样品标准品或含目的外源基因的质粒作为标准品来建立标准曲线,继而得到样品的拷贝数,但是,标准品的获得过程较为繁琐,且标准曲线的建立对标准品DNA的质量、引物和探针等因素的要求也较高25]。本研究建立的转基因糜子外源基因插人拷贝数qPCR检测方法,无需建立标准曲线,只需以已知插人拷贝数的转基因样品为参照,采用相对定量的方法来初步计算样品中外源基因的插人拷贝数,简化了实验方法,节省了实验成本和时间。

dPCR技术是在qPCR技术基础上的一次技术新突破,是对核酸分子的绝对定量。dPCR与qPCR相比,准确度、灵敏度以及特异性都得到了进一步提升。dPCR也可用于检测外源基因在受体植物基因组中的插人拷贝数,且已被应用在多种转基因植物中,如水稻、玉米、番茄、柑橘、小麦和马铃薯[26]。Narancio等[27]使用qPCR 和 dPCR 检测外源基因在白三叶中的插人拷贝数,结果发现dPCR具有更高的准确性。
本研究也首次成功建立了利用dPCR检测转基因糜子植株中外源基因插人拷贝数的方法,由于dPCR不受PCR扩增效率影响,对外源基因拷贝数的检测有更高的准确性,但其仪器设备不易获得,且成本较高,不适用于大量样本的检测,在有条件的情况下可使用该方法。相比之下,qPCR成本较为经济,操作简单,可以批量检测,特别适合用于从大量转基因植株中初步筛选出以低拷贝插人到受体基因组的转基因植株;随后,可利用Southern杂交技术对初步筛选的植株进行验证,极大地节省了时间和成本。
3.2结论
本研究利用dPCR方法测定了9个转基因糜子样品中的内参基因SDI和外源基因 Hy g 的拷贝数,根据 Hy g 与 S D I 的拷贝数比值得到了9个样品的外源基因插人拷贝数;其中,有8个样品为双拷贝插入,1个样品为单拷贝插入。根据dPCR检测的结果,选择一株双拷贝插人的转基因糜子样品作为qPCR的参照样本,建立了利用qPCR快速检测转基因糜子植株中 _{H y g} 基因的相对拷贝数的方法。23份转基因糜子样品的检测结果显示,3份样品是单拷贝插人的转基因植株,7份样品是双拷贝插入的转基因植株,其余样品均为三拷贝及以上插入的转基因植株。对其中的1份单拷贝样品和3份双拷贝样品也进行了dPCR检测,结果与qPCR检测结果完全一致,说明gPCR法检测外源基因低拷贝插入时的准确度较高,可以满足转基因糜子育种研究中低拷贝株系初步筛选的要求。本研究建立的利用dPCR 和qPCR技术检测转基因糜子植株中外源基因插人拷贝数的方法,简单快速,样本用量少,均可作为转基因糜子育种中外源基因插人拷贝数检测的新方法。
参考文献
[1]Bhat S, Nandini C, et al. Proso millet (Panicum miliaceum L.)-a climate resilient crop for food and nutritional security: A Review[J]. Environment Conservation Journal, 2019,
20(3): 113-124. [2] Flajsman M, Stajner N, Kocjan Acko D. Genetic diversity and agronomic performance of Slovenianlandraces of proso millet (Panicum miliaceum L.)[J]. Turkish Journal of Botany,
2019, 43(2): 185-195.
[3]Calamai A, Masoni A, Marini L, et al. Evaluation of the agronomic traits of 80 accessions of proso millet (Panicum miliaceum L.) under Mediterranean pedoclimatic conditions[J]. Agriculture, 2020, 10(12): 578.
[4]Lu H Y, Zhang JP, Liu K B, et al. Earliest domestication of common millet (Panicum miliaceum) in East Asia extended to 10, 000 years ago[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(18): 7 367-7 372.
[5] Hunt HV, Badakshi F, Romanova O, et al. Reticulate evolution in Panicum (Poaceae): The origin of tetraploid broomcorn millet, P. miliaceum[J] Journal of Experimental Botany, 2014, 65(12): 3 165-3 175.
[6]杨清华,王洪露,冯佰利.糜子品质研究进展与展望[ 植物学报,2023,58(1):22-33.
[7]李锦华,朱新强,余成群,等.西藏高海拔河谷农区糜子 种植前景展望[].中国草食动物科学,2023,43(4):54-58.
[8]Zou C S, Li L T, Miki D, et al. The genome of broomcorn millet[J]. Nature Communications, 2019, 10(1): 436.
[9]杨天育.糜子分子遗传研究进展与展望[]].寒旱农业科学, 2022(10): 32-36.
[10]盘婉向,刘天鹏,何继红,等.糜子(Panicumn miliaceum L.)全基因组Y4BBY基因家族鉴定与高渗溶液胁迫下表 达特征[].基因组学与应用生物学,2022,41(5):1067- 1 078.
[11] Liu Y, Cheng Z X, Chen W Y, et al. Establishment of genome-editing system and assembly of a near-complete genome in broomcorn millet[J]. Journal of Integrative Plant Biology, 2024, 66(8): 1 688-1 702.
[12]Lu Q, Zhao H N, Zhang Z Q, et al. Genomic variation in weedy and cultivated broomcorn millet accessions uncovers the genetic architecture of agronomic traits[J]. Nature Genetics, 2024, 56(5): 1 006-1 017.
[13] Collier R, Dasgupta K, Xing Y P, et al. Accurate measurement of transgene copy number in crop plants using droplet digital PCR[J]. The Plant Journal, 2017, 90(5): 1 014-1 025.
[14]Stam M. Review article: The silence of genes in transgenic plants[J]. Annals of Botany, 1997, 79(1): 3-12.
[15] Mason G, Provero P, Vaira A M, et al. Estimating the number of integrations in transformed plants by quantitative real-time PCR[J].BMC Biotechnology, 2002, 2: 20.
[16]谢秀菊,夏启玉,刘帅,等.数字PCR和荧光定量PCR检 测转基因番木瓜中外源基因拷贝数方法的建立及其应用 [].热带作物学报,2024,45(4):663-673.
[17]杨立桃,赵志辉,丁嘉羽,等.利用实时荧光定量PCR方 法分析转基因水稻外源基因拷贝数[.中国食品卫生杂 志,2005, 17(2): 140-144.
[18]王育花,赵森,陈芬,等.利用实时荧光定量PCR 法检测 转基因水稻外源基因拷贝数的研究[川.生命科学研究,
2007, 11(4): 301-305. [19]魏毅东,毛小辉,何炜,等.利用多重荧光定量PCR检测 转基因水稻外源基因拷贝数[J.农业生物技术学报,2017,
25(12): 2 072-2 078. [20]闫喜中,张锐,孟志刚,等.实时荧光定量PCR 法检测抗 虫棉Bt基因拷贝数方法的建立[J].安徽农业科学,2008,
36(26): 11 376-11 377. [21]石建斌,杨永智,王舰.马铃薯T-DNA插人拷贝数的检测 及对农艺性状的影响研究[川.中国农业大学学报,2015,
20(6): 68-75. [22]许兰珍,何永睿,雷天刚,等.转基因柑橘外源基因拷贝 数的实时荧光定量PCR检测[J].园艺学报,2016,43(6):
1 186-1 194. [23]任爽,朱鸿亮.Taqman定量PCR技术检测基因编辑番茄 中外源基因拷贝数体系的建立川.中国生物工程杂志,
2017, 37(10): 72-80.
[24]铁原毓,文军琴,田洁.转As6G-FFT基因烟草的阳性鉴 定及基因拷贝数测定[J].福建农业学报,2022,37(5): 592-599.
[25]Cankar K, Stebih D, Dreo T,et al. Critical points of DNA quantification by real-time PCR—effects of DNA extraction method and sample matrix on quantification of genetically modified organisms[].BMC Biotechnology,2006,6:37.
[26] Hindson B J, Ness K D, Masquelier D A, et al. High-throughput droplet digital PCR system for absolute quantitation of DNA copy number[J]. Analytical Chemistry, 2011, 83(22): 8 604-8 610.
[27]Narancio R, John U, Mason J, et al. Digital PCR (dPCR) and qPCRmediated determination of transgene copy number in the forage legume white clover (Trifolium repens)[J]. Molecular Biology Reports, 2021, 48(4): 3 069-3 077.
激发子蛋白SbES高密度重组表达和粉剂研发
杨扬」陈奕鹏」王茂存²章淑艳²刘先宝黄贵修1
(1.中国热带农业科学院环境与植物保护研究所/农业农村部热带作物有害生物综合治理重点实验室/海南省热带农业有害生物监测与控制重点实验室/海南省热带作物病虫害生物防治工程技术研究中心海南海口571101:2河业少微生物研空所右限八司河北促宝071051)
摘要激发子蛋白SbES 是帚枝霉属内生真菌HND5产生的一个外泌丝氨酸蛋白酶,可有效诱导多种植物产生抗病性,具有可商品化开发为植物蛋白农药的潜力。为建立该蛋白的高密度发酵及粉剂制备工艺,利用已构建好的SbES蛋白毕赤酵母异源表达菌株Pichia pastorisX-33(pRICZA:SbES),对该蛋白的高密度发酵条件、菌体破碎条件及适宜喷雾干燥条件进行优化。结果表明,在测试范围内,SbES蛋白毕赤酵母异源表达菌株在 \mathsf{p H} 6.5 、温度 28 °C 和菌体浓度180~g/L 条件下诱导 108~h ,可获得最大表达量;目标异源表达菌株 15% 菌体浓度,在 300~W 功率下超声 40\;min ,可获得最大的SbES蛋白量;SbES蛋白最适的助干剂为麦芽糊精,在 5% 麦芽糊精,干燥塔出风口温度为 140 °C 的条件下干燥,SbES 蛋白可保存最大的酶活性。本研究结果为激发子蛋白SbES 的商品化开发提供了研究基础。
关键词激发子蛋白;异源表达;高密度发酵;粉剂制备;植物蛋白农药
High-cell-density Recombinant Expression and Powder Development of the Elicitor Protein SbES
YANG Yang' CHEN Yipeng' WANG Maocun² ZHANG Shuyan² LIU Xianbao' HUANG Guixiu'
(1. Environment and Plant Protection Institute, Chinese Academy of Tropical Agricultural Sciences/Key Laboratory of Comprehensive Management of Tropical Crop Pests, Ministry of Agriculture and Rural Affairs/Hainan Key Laboratory for Monitoring and
Control of Tropical Agricultural Pests/Hainan Engineering Research Center for Biological Control of Tropical Crops Diseases and Insect Pests, Haikou, Hainan 571101, China; 2. Hebei Research Institute of Microbiology Company Limited, Baoding,Hebei 071051,China)
Abstract The elicitor protein SbES is a secreted serine protease found in the endophytic fungus Sarocladium sp.HND5. This protein can effectively induce disease resistance in a variety of plants and has the potential to be commercialized as a plant protein pesticide. In this study, we used the heterologous expression strain Pichia pastoris X-33 (pRICZA:SbES) to establish a high-density fermentation and powder preparation process for the protein. The results revealed that Pichia pastoris X-33 (pRICZA:SbES) had the maximum SbES expression amount under the conditions at pH 6.5, 28 °C , and a cell concentration of 180~g/L .The heterologous expression strain could be disruptedwellvia 300~W ultrasonic disruption for 40\ min with a cell density of 15% . The most suitable drying agent for the SbES protein was maltodextrin. The maximum enzyme activity of the SbES protein was preserved after drying at 140 °C with 5% maltodextrin.The results of this study provide a research basis for thecommercialdevelopmentoftheelicitorproteinSbES.
Keywords elicitor protein; heterologous expression; high-cell-density fermentation; protein powder preparation; plant proteinpesticide
植物病原菌的某些外泌水解酶或无毒蛋白可被植物识别,并将信号逐级传导至细胞内部,激发植物的免疫反应,这类物质被统称为激发子蛋白川。此类蛋白可通过与植物相关受体结合,激发植物产生活性氧爆发、朕质积累等抗病反应,并通过信号传导,诱发程序性细胞死亡和过敏性反应,从而提高植物抗病性。目前,已经报道的植物激发子蛋白主要有来自细菌的Harpin 蛋白²,Alternaria tenuissima 中的PeaT1和 Hrip13],Verti-cillium dahliae 中的 VdAL 和 PevD14],Pucciniatriticina 中的 Pt18906^{[5]} ,Botrytis cinerea 中的BcGs1,以及 Acremonium strictum 分泌的胞外蛋白酶AsES等[]。
激发子蛋白SbES是帚枝霉属内生真菌HND5产生的一个外泌丝氨酸蛋白酶,前期研究发现:该蛋白可有效激发辣椒叶片活性氧暴发、微过敏反应和肼质积累等抗病性反应;能有效提高辣椒叶片中与活性氧暴发、过敏性反应、肼质合成、植保素合成等抗病反应相关的基因及水杨酸、茉莉酸和乙烯信号传导关键基因的表达。以 0.1\ mg/mL 浓度的SbES蛋白溶液喷施辣椒叶片,可有效提高辣椒对棒孢叶斑病的抗性"。同时,低浓度SbES重组蛋白处理也可有效提高木薯对细菌性枯萎病的抗性”。相关结果表明,该蛋白具有可商品化开发为植物蛋白农药的潜力,但并未进行大规模生产工艺的相关研究。
毕赤酵母(Pichiapastoris)是一种成熟且广泛使用的蛋白表达平台,可高效表达源自真核生物的目的蛋白。该系统具有细胞生长快、翻译后修饰系统完整、重组蛋白纯度高和遗传稳定性高等优势,广泛用于真菌蛋白的规模化生产。本研究前期利用毕赤酵母X-33菌株构建了激发子蛋白SbES的异源表达菌株PichiapastorisX-33(pRICZA::SbES),并通过摇瓶培养获得了SbES的重组蛋白。但通过摇瓶发酵获得的重组蛋白量少,无法进行规模化生产,限制了SbES 蛋白农药的开发和利用。本研究拟探究初始诱导菌体浓度、诱导温度和pH对异源表达菌株PichiapastorisX-33(pRICZA::SbES)高密度发酵水平的影响,通过优化超声功率、菌体浓度和超声时间,获得异源表达菌株最适菌体破碎条件;筛选合适的SbES蛋白助干剂,优化助干剂浓度及喷雾干燥温度,获得适合SbES蛋白的粉剂制备工艺,为后期的商品化开发及工业化生产奠定基础。
1 材料与方法
1.1材料
1.1.1菌株激发子蛋白SbES异源表达菌株PichiapastorisX-33(pRICZA::SbES),以及空白对照菌株P.pastorisX-33(pRICZA)由本实验室构建和保藏。
1.1.2仪器格瑞生物GR-BL10联补料控制系统+Kinco10联灭菌玻璃发酵系统;苏净安泰SW-CJ-1FD无菌超净工作台;SCIENTZ-950E超声波细胞破碎仪;JOYN-8000T乔跃实验室小型喷雾干燥机;INESA-L3S可见分光光度计;君意JY60OHE蛋白电泳仪;天能5200凝胶成像系统。1.1.3试剂博来霉素,Anti-His Tag Monoclonalantibody,AffiniPure Goat Anti-Mouse IgG,ECLPLUS超敏发光液,索莱宝科技(北京)有限公司;微生物丝氨酸蛋白酶酶活检测试剂盒,酶免(江苏)有限公司;PVDF膜,碧云天生物技术公司(上海);M5PrecisePrestainedProteinLadder10-180\;kDa) ,聚合美生物科技有限公司(北京);His-Trap亲和柱,Sigma-Aldrich;其他试剂均为国产分析纯。
1.1.4主要培养基LB、YPD、MD、BMGY与BMMY配方参照Invitrogen公司的《毕赤酵母表达操作手册》。PTM1培养基成分为五水合硫酸铜6.00~g 、碘化钠 \mathbf{0.08~g} 、一水硫酸锰 3.00~g 、钼酸钠 0.20~g 、硼酸 _{0.02up{g}} 、氯化钴 0.50~g 、硫酸锌20.00\ g 、七水硫酸亚铁 65.00 bf{g} 、VH(生物素)0.20~g 、硫酸 5.00\;mL ,用蒸馏水定容至 1\ 000\ mL ,过滤灭菌,室温保存;BSM(高密度发酵培养基)成分为 H_{3}PO_{4} 26.70\;~mL~ 、 CaSO_{4} 0.93~g~ 、 K_{2}SO_{4} 18.20~g 、 MgSO_{4{*}7H_{2}O\ 14.90\ g} KOH\ 4.13\ g 、甘油 40{~g~} C 31.67~mL )、PTM1 4.35~mL ,调整 \mathsf{p H} 至5.00,蒸馏水定容至 1~L~ ,搅拌溶解,转移至发酵罐后以 121 °C 灭菌;甘油培养基成分,将 50% 甘油灭菌后,添加 1.20% 过滤除菌的PTM1微量元素溶液;甲醇培养基成分,在甲醇中添加 1.20% 过滤除菌的PTM1微量元素溶液; 10{x}PBS Buffer成分为氯化钠 40{~g~} ,氯化钾 _{~l~g~} ,磷酸氢二钠 7.10\;g ,磷酸二氢钾 1.35~g~ ,蒸馏水定容至 500~mL
1.2方法
1.2.1P.pastorisX-33(pRICZA::SbES)发酵罐高密度发酵及条件优化挑取平板中的 P. pas-torisX-33(pRICZA::SbES)单菌落至装有 5mL YPD ( 1%_{0} 博来霉素)灭菌试管中,接种,过夜培养;按照 10% 接种量将试管菌体转移至装有100~mL BMGY培养基的 500~mL 摇瓶中培养 6~ 7 h ( O D_{600} 4~5\ )_{\circ} 配制5LBSM培养基,放入 10~L 生物发酵罐中,以 121 °C 灭菌 20\ min ;待冷却至30 °C 左右过滤,加入 4.35~mL/L PTM1微量元素培养基,调整转速至 700~r/min ,用氨水调整培养基pH至5.00,接种 10% 摇瓶种子液, 30 °C 培养。发酵溶氧升高至 100% 时,进入甘油补料阶段,每升发酵液以 18.15 \mL/h 速度流加 50% 甘油 4 ~h~ ,湿重到达 180~220~g/L 且发酵溶氧不再升高时,说明甘油消耗完毕,进人甲醇诱导阶段。调整诱导温度(28、30和 32 °C \mathsf{p H} (4.5、5.5和6.5)和初始诱导菌体湿重(160、180和 200~g/L ),开始流加甲醇培养基,初始为每升发酵液 1.50\:~mL/h 流加 10~h ;之后调整流速为 2.00\;mL/h ,继续流加甲醇 48 h ,期间每 12 h 取样1次,用于丝氨酸蛋白酶酶活测定;待诱导 70 h 后,调整流速为 4.00\;mL/h ,保持至发酵结束,诱导时间为 96~120\;h_{c} 在整体发酵期间,每 24~h 添加少量VC以保持菌体活性并维持溶氧值在 20% 以上。
取 1 rm{m L} 发酵菌液样品,离心收集菌体后使用, 1\ mL\pH\ 6.5 PBS缓冲液悬浮,超声波破碎后,10\ 000\ r/min 离心 10\;min ,取上清,使用酶联免疫试剂盒检测丝氨酸蛋白酶酶活。根据酶活的大小,确定最适高密度发酵条件。
1.2.2P.pastorisX-33(pRICZA::SbES)菌体破碎条件优化SbES蛋白为胞内表达蛋白,进一步对菌体破碎条件进行优化。将发酵后的酵母菌体离心,弃上清,菌体沉淀用PBS缓冲液重悬,按照初始拟定的破碎条件进行细胞破碎:功率300~W , 15% 菌体浓度,超声5s暂停 5\;s ,冰浴条件下超声 40\;min ,离心,留取上清进行酶活检测。通过改变超声功率(200、300、400、500W)、菌体浓度( 5% 、 10% 、 15% 、 20% )时间(20、30、40、 50\;min ),根据破碎后上清丝氨酸蛋白酶酶活确定最佳超声破碎条件。
1.2.3SbES蛋白粉剂制备方法研发将超声破碎后的菌体以 10 000 r/min 离心 10 \min ,留取上清液;在上清液中分别加人 2% 的麦芽糊精、可溶性淀粉、 β -环糊精、脱脂奶粉,搅拌均匀后,以进风温度 140 °C 、蠕动泵流速 18~r/min 进行喷雾干燥;收集粉剂并加水溶解至原体积,测量丝氨酸蛋白酶的酶活力,筛选最优干燥助剂。在上清液中加人一定比例的助干剂,分别选择一系列进风温度110、120、130、140、 150 °C ,以相同的出风温度、流速 18 \Deltar/min 进行喷雾干燥,加水溶解至原体积后测量丝氨酸蛋白酶的酶活力,根据酶活大小筛选最优进风温度。
1.2.4丝氨酸酶活力检测按照微生物丝氨酸蛋白酶(PRSS)酶联免疫分析(酶免)试剂盒说明书进行。
1.2.5P.pastorisX-33(pRICZA)丝氨酸蛋白酶酶活检测按照1.2.1的方法进行 P. pastoris X-33(pRICZA)菌株的高密度发酵。加入甲醇诱导后,每隔 24~h 取 1~mL 发酵菌液样品,离心收集菌体,使用 ~1~mL~pH~6.5~ PBS缓冲液悬浮,超声波破碎后,以 10 \ 000 \ r/min 离心 10 \min ,取上清,使用酶联免疫试剂盒检测丝氨酸蛋白酶酶活。
1.2.6SbES重组蛋白纯化参考前期实验方法[8进行SbES蛋白的纯化。
1.2.7WesternBlot实验方法(1)SDS-PAGE电泳检测:吸取每管上清溶液 20~\upmuL ,与 5\ \upmuL\ 5x 蛋白上样buffer混匀,水浴加热煮沸 10 \min ,冰浴 5~min ,以 4 °C 、 12\ 000\ r/min 离心 10\ min ;吸取上清样品 15~\upmuL 上样,利用蛋白Marker 10~\upmuL SDS-PAGE进行电泳检测。(2)提前用甲醇浸泡激活PVDF膜,按照正极电源一海绵垫一厚滤纸一PVDF膜一蛋白凝胶一厚滤纸一海绵垫一负极电源(黑胶红膜)的顺序完成“三明治"转膜装置的组装,放入电泳槽中,冰浴条件下以 100~mA 恒流转膜 70\ min 。(3)取出PVDF膜放人孵育盒中,加入 3%BSA 封闭液至没过蛋白膜,室温缓慢震荡 4 h 。(4)按 1:5000 (体积比)在封闭液中加人一抗,室温缓慢震荡 1 ~h~ 。(5)弃掉封闭液,加人TBST溶液清洗蛋白膜,重复3次,每次清洗震荡 10\;min 。(6) 1:2\ 000 (体积比)加人二抗,室温缓慢震荡 ^{1~h~} 。(7)将发光液A、B按照1:1 (体积比)比例混合后,避光孵育整个PVDF膜,利用CCD冷冻低温照相机显影。
2 结果与分析
2.1发酵罐条件下SbES的诱导表达及丝氨酸 蛋白酶酶活检测
结果显示,发酵罐培养条件下,SDS-PAGE及WesternBlot(His标签检测)均能检测得到34kDa大小的特异片段(图1)。该结果同前期摇瓶结果一致,表明发酵罐高密度培养条件下, P. pastorisX-33(pRICZA::SbES)菌株中的SbES蛋白得到了有效表达。
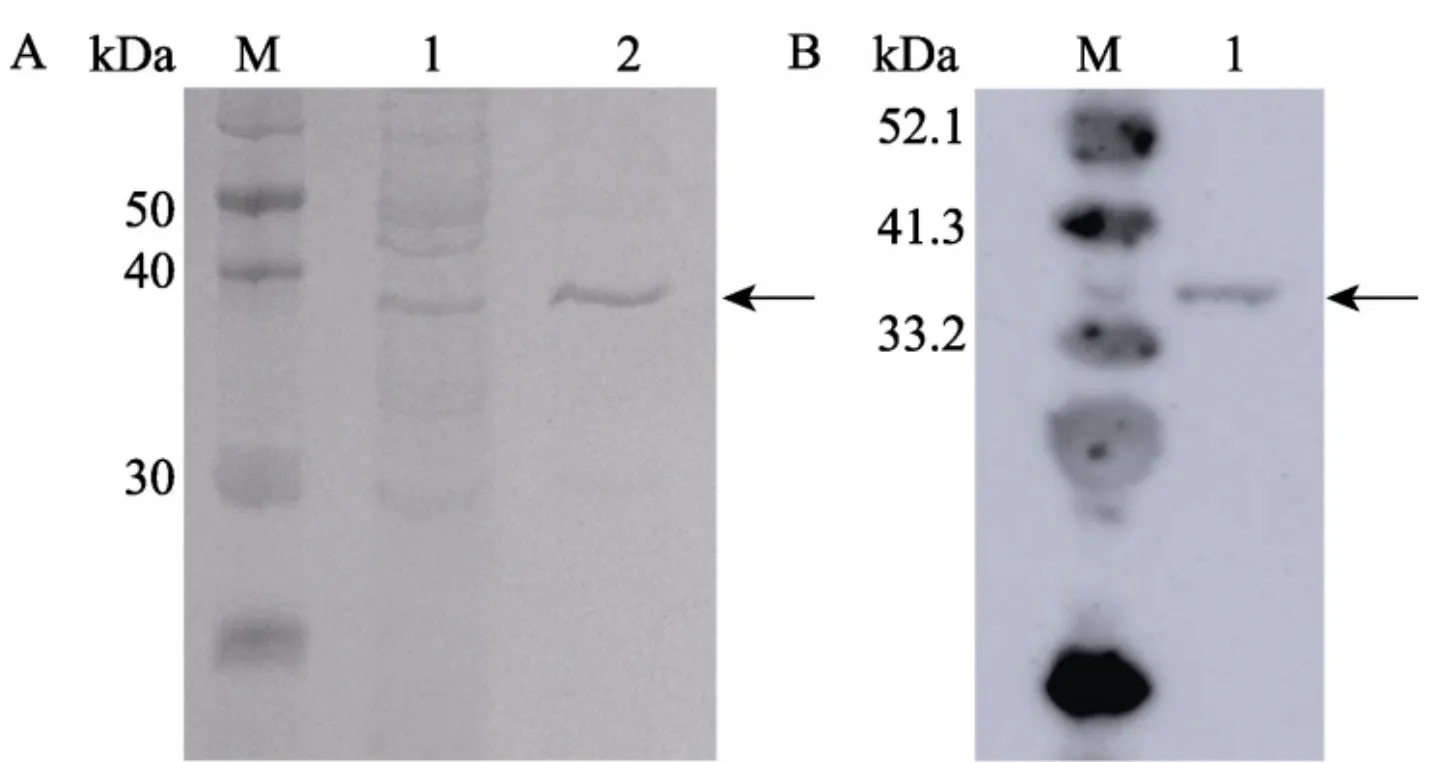
图1SbES蛋白异源表达检测
根据氨基酸残基序列BLAST分析,SbES蛋白具有丝氨酸蛋白酶酶活。利用酶联免疫丝氨酸蛋白酶酶活检测试剂盒对纯化的SbES重组蛋白进行酶活检测发现,未使用甲醇诱导时 P_{*} pastorisX-33(pRICZA::SbES)菌体上清丝氨酸蛋白酶酶活在 100~200\ IU/L ,使用甲醇诱导表达后可达到500 rm{IU}/L 。同时,对转入空白pRICZA载体的 P. pastorisX-33菌株[P.pastoris X-33(pRICZA)]高密度发酵条件下的丝氨酸蛋白酶酶活进行检测发现,在高密度发酵条件下, P_{\star} pastoris X-33(pRICZA)菌体上清丝氨酸蛋白酶酶活在25IU/L左右(图2),仅为异源表达菌株 P. pastorisX-33(pRICZA::SbES)的 5% ,表明异源表达菌株 P. pastorisX-33(pRICZA::SbES)丝氨酸蛋白酶酶活主要来自于SbES蛋白,可以通过检测丝氨酸蛋白酶酶活检测SbES蛋白的表达量。
2.2不同诱导 \mathbf{pH} 对SbES异源表达的影响
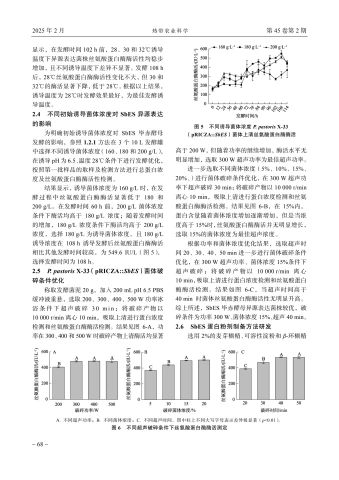
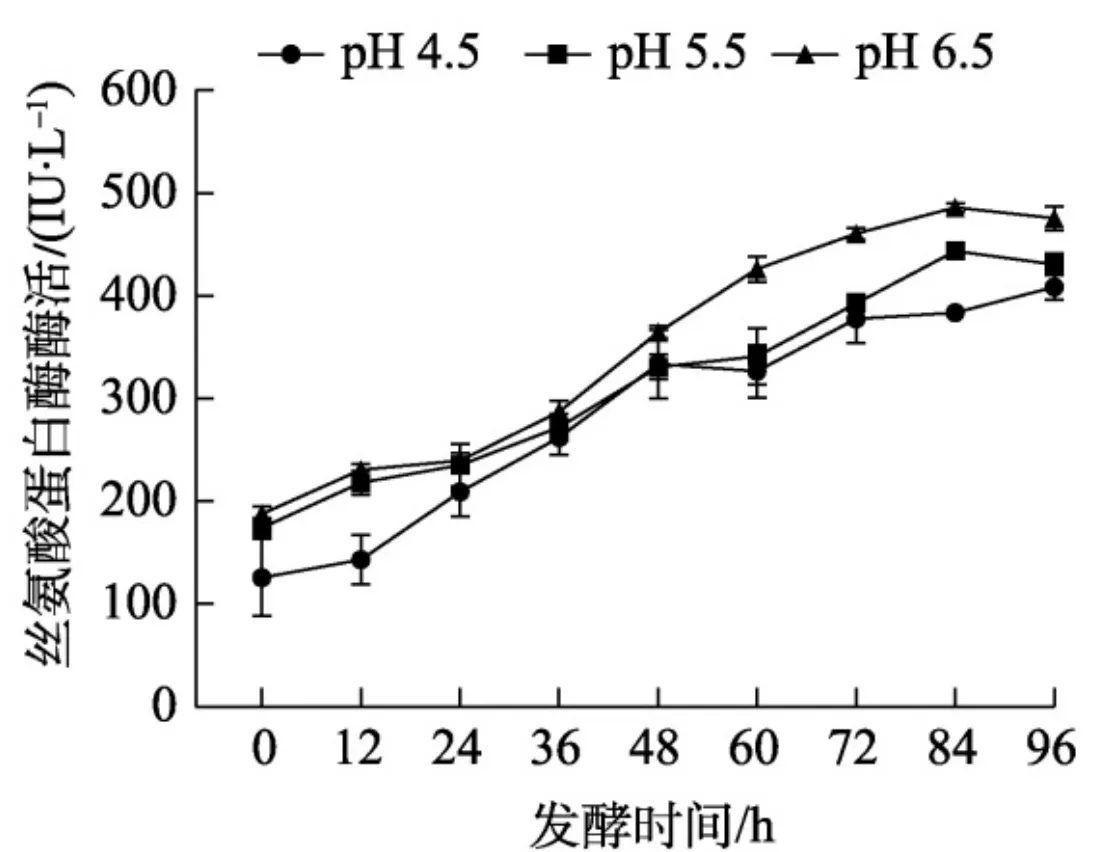
首先对SbES发酵的pH条件进行优化,参照1.2.1方法,在3个 10 ~L~ 发酵罐中选择不同诱导pH(4.5、5.5、6.5)进行发酵条件优化和丝氨酸蛋白酶酶活性检测。结果显示,菌体上清丝氨酸蛋白酶酶活随着诱导时间的增加而提高, 48 ~h~ 前不同诱导pH发酵条件下活性差别小, 48 h 后pH6.5发酵条件下活性显著高于 \mathsf{p H}\ 5.5 、4.5,选择\mathsf{p H} 6.5 为最佳发酵pH(图3)。
2.3不同诱导温度对SbES异源表达的影响
为明确温度对SbES异源表达的影响,参照1.2.1方法,在3个 10 ~L~ 发酵罐中选择不同诱导温度(28、30、 32 °C ),在诱导pH为6.5条件下进行发酵优化,按照第一批样品的取样及检测方法进行丝氨酸蛋白酶酶活性检测。结果(图4)


显示,在发酵时间 102~h 前,28、30和 32 °C 诱导温度下异源表达菌株丝氨酸蛋白酶酶活性均稳步增加,且不同诱导温度下差异不显著。发酵 108~h 后, 28 °C 丝氨酸蛋白酶酶活性变化不大,但30和32 °C 的酶活显著下降,低于 28 °C 。根据以上结果,诱导温度为 28 °C 时发酵效果最好,为最佳发酵诱导温度。
2.4不同初始诱导菌体浓度对SbES异源表达的影响
为明确初始诱导菌体浓度对SbES毕赤酵母发酵的影响,参照1.2.1方法在3个 10~L 发酵罐中选择不同诱导菌体浓度(160、180和 200~g/L 在诱导pH为6.5、温度 28 °C 条件下进行发酵优化,按照第一批样品的取样及检测方法进行总蛋白浓度及丝氨酸蛋白酶酶活性检测。
结果显示,诱导菌体浓度为 160~g/L 时,在发酵过程中丝氨酸蛋白酶酶活显著低于180和200~g/L 。在发酵时间 60~h 前, 200~g/L 菌体浓度条件下酶活均高于 180~g/L 浓度;随着发酵时间的增加, 180~g/L 浓度条件下酶活均高于 200~g/L 浓度,选择 180~g/L 为诱导菌体浓度。且 180~g/L 诱导浓度在 108~h 诱导发酵后丝氨酸蛋白酶酶活相比其他发酵时间较高,为 549.6\ IU/L (图5),选择发酵时间为 108~h
2.5PpastorisX-33(pRICZA:SbES)菌体破碎条件优化
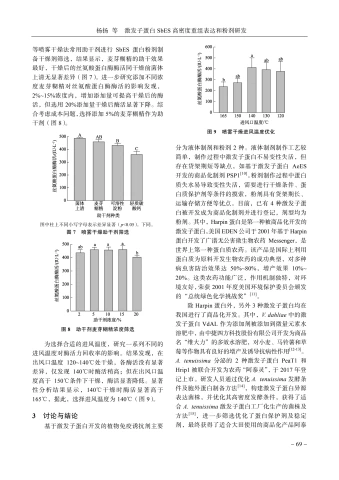
称取发酵菌泥 _{20up{g}} ,加人 200~mL~pH~6.5 PBS缓冲液重悬,选取200、300、400、500W功率冰浴条件下超声破碎 3\ 0\ ~m~i~n~ ;将破碎产物以10\ 000\ r/min 离心 10\ min ,吸取上清进行蛋白浓度检测和丝氨酸蛋白酶酶活检测。结果见图6-A,功率在300、400和 500 W 时破碎产物上清酶活均显著高于 200~W ,但随着功率的继续增加,酶活水平无明显增加,选取 300\;W 超声功率为最佳超声功率。

进一步选取不同菌体浓度( 5% 10% 、 15% 20% 、)进行菌体破碎条件优化,在 300~W 超声功率下超声破碎 30\;min ;将破碎产物以 10\ 000\ r/min 离心 10\ min ,吸取上清进行蛋白浓度检测和丝氨酸蛋白酶酶活检测。结果见图6-B,在 15% 内,蛋白含量随着菌体浓度增加逐渐增加,但是当浓度高于 15% 时,丝氨酸蛋白酶酶活并无明显增长,选取 15% 的菌体浓度为最佳超声浓度。
根据功率和菌体浓度优化结果,选取超声时间20、30、40、 50\;min 进一步进行菌体破碎条件优化,在 300\;W 超声功率、菌体浓度 15% 条件下超声破碎;将破碎产物以 10\ 000\ r/min 离心10\;min ,吸取上清进行蛋白浓度检测和丝氨酸蛋白酶酶活检测。结果如图6-C,当超声时间高于40\;min 时菌体丝氨酸蛋白酶酶活性无明显升高。综上所述,SbES毕赤酵母异源表达菌株较优,破碎条件为功率 300\;W 、菌体浓度 15% 、超声 40\;{min} 。
2.6SbES蛋白粉剂制备方法研发
选用 2% 的麦芽糊精、可溶性淀粉和 β 环糊精等喷雾干燥法常用助干剂进行SbES蛋白粉剂制备干燥剂筛选,结果显示,麦芽糊精的助干效果最好,干燥后的丝氨酸蛋白酶酶活同干燥前菌体上清无显著差异(图7)。进一步研究添加不同浓度麦芽糊精对丝氨酸蛋白酶酶活的影响发现,2%~15% 浓度内,增加添加量可提高干燥后的酶活,但选用 20% 添加量干燥后酶活显著下降。综合考虑成本问题,选择添加 5% 的麦芽糊精作为助干剂(图8)。
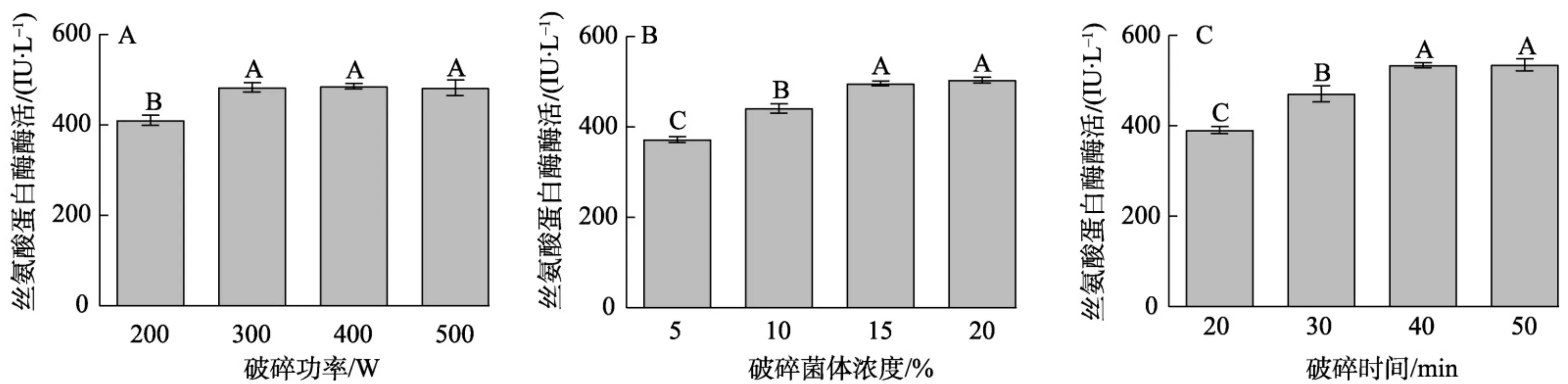
图6不同超声破碎条件下丝氨酸蛋白酶酶活测定

图7喷雾干燥助干剂筛选

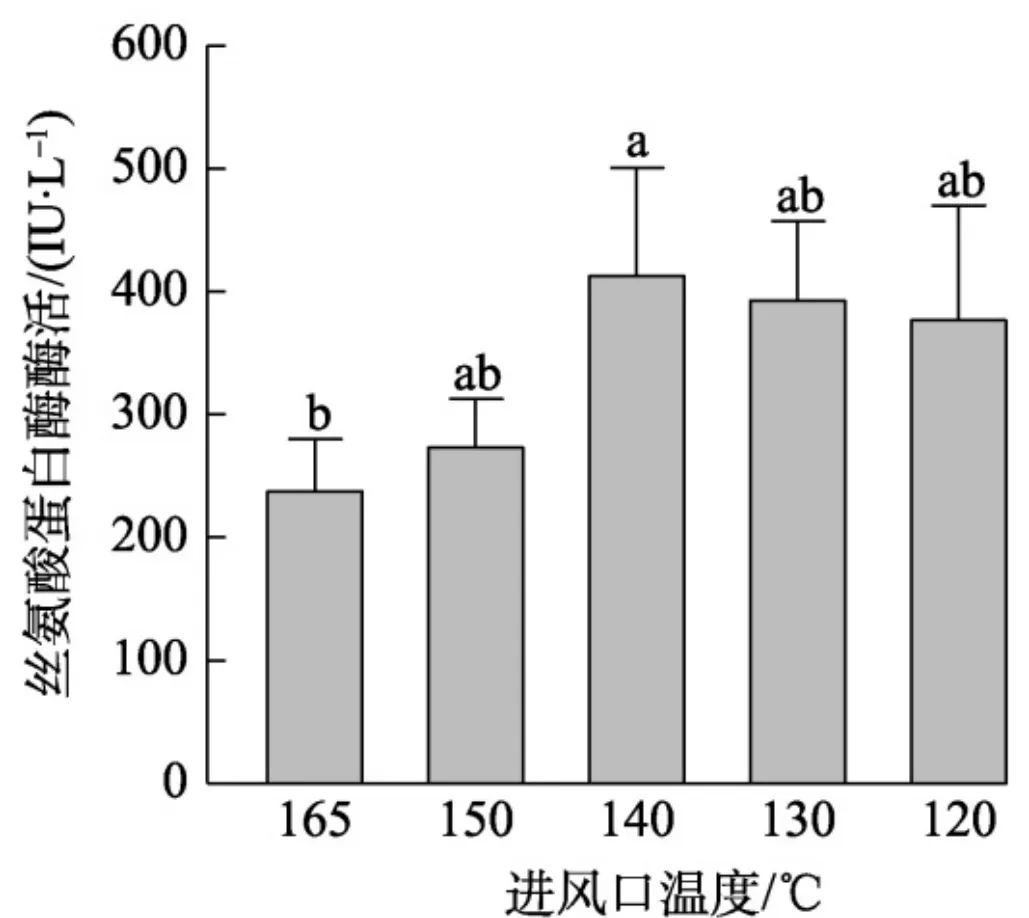
为选择合适的进风温度,研究一系列不同的进风温度对酶活力回收率的影响。结果发现,在出风口温度 120~140°C 处干燥,各酶活没有显著差异,仅发现 140 °C 时酶活稍高;但在出风口温度高于 150 °C 条件下干燥,酶活显著降低。显著性分析结果显示, 140 °C 干燥时酶活显著高于165 °C ,据此,选择进风温度为 140 °C (图9)。
3 讨论与结论
基于激发子蛋白开发的植物免疫诱抗剂主要分为液体制剂和粉剂2种。液体制剂制作工艺较简单,制作过程中激发子蛋白不易变性失活,但存在货架期短等缺点,如基于激发子蛋白AeES开发的商品化制剂PSP1。粉剂制作过程中蛋白质失水易导致变性失活,需要进行干燥条件、蛋白质保护剂等条件的摸索,粉剂具有货架期长、运输存储方便等优点。目前,已有4种激发子蛋白被开发成为商品化制剂并进行登记,剂型均为粉剂。其中,Harpin蛋白是第一种被商品化开发的激发子蛋白,美国EDEN公司于2001年基于Harpin蛋白开发了广谱无公害微生物农药Messenger,是世界上第一种蛋白质农药。该产品是国际上利用蛋白质为原料开发生物农药的成功典型,对多种病虫害防治效果达 50%~80% ,增产效果 10%~ 20% 。这类农药功能广泛,作用机制独特,对环境友好,荣获2001年度美国环境保护委员会颁发的“总统绿色化学挑战奖”[1]。
除Harpin蛋白外,另外3种激发子蛋白均在我国进行了商品化开发。其中,Vdahliae中的激发子蛋白VdAL作为添加剂被添加到微量元素水溶肥中,由中捷四方科技股份有限公司开发为商品名“维大力”的多效水溶肥,对小麦、马铃薯和草莓等作物具有良好的增产及诱导抗病性作用[12-13]。A.tenuissima分泌的2种激发子蛋白PeaT1和Hrip1被联合开发为农药“阿泰灵”,于2017年登记上市。研发人员通过优化A.tenuissima发酵条件及胞外蛋白制备方法[4],构建激发子蛋白异源表达菌株,并优化其高密度发酵条件,获得了适合A.tenuissima激发子蛋白工厂化生产的菌株及方法[15],进一步筛选优化了蛋白保护剂及稳定剂,最终获得了适合大田使用的商品化产品阿泰灵,可有效提高水稻、葡萄和番茄对纹枯病、霜霉病和黄化曲叶病毒病的抗性[16-1]
SbES是从内生帚枝霉属真菌HND5菌株中鉴定得到的一个激发子蛋白,可诱导辣椒产生对多主棒孢霉菌的抗性,并能激发辣椒和木薯叶片产生活性氧暴发、微过敏反应、朕眠质积累等抗病反应,诱导抗病相关基因的表达,具有开发为植物诱抗剂的潜力[-]。前期研究构建了SbES 蛋白的毕赤酵母异源表达菌株,但摇瓶培养条件下目的蛋白产量小,限制了相关产品的研发。毕赤酵母非常适合发酵生长,在发酵过程中可以实现细胞的高密度生长,是毕赤酵母大规模生产外源蛋白的重要策略[]。毕赤酵母的生长 pH 较为宽泛 pH~2.8~8.0 ),但诱导环境的pH变化会影响菌体的生长状态、细胞通透性,直接影响目的蛋白表达水平与活性。本研究结果表明,目的蛋白异源表达菌株 P_{*} pastoris X-33(pRICZA:SbES)在 \mathsf{p H} 6.5 诱导条件下的丝氨酸酶活显著高于 pH4.5、5.5,说明在验证范围内,6.5是最适诱导 {\mathfrak{p H}}
温度是影响微生物发酵、调控质量的重要外因,一方面对细胞的生长有重要影响,另一方面对代谢调控关键酶的活性也有决定性的影响[1]。本研究对不同诱导温度下目标菌株丝氨酸酶活进行检测,发现 28 °C 条件下诱导的酶活性明显超过30和 32 °C ,为最适诱导温度。进一步的诱导菌体浓度优化实验结果表明,诱导菌体浓度为 180~g/L 时发酵效果较好,在进行 108 ~h~ 诱导发酵后丝氨酸蛋白酶酶活为 549.6~{IU/L} 。通过酶活的测定结果可以看出,并不是接种量越高,最终发酵完成的酶活力就越高。接种量过低,发酵培养时间过长,发酵过程中动力的消耗会增加,并且有染杂菌风险;接种量过高会导致发酵前期菌体生长过于旺盛,代谢产物增多,造成菌体的早衰。此外,接种量过高也会消耗大量的营养物质,从而影响目的蛋白的合成。汪飞等2利用毕赤酵母高密度表达 β* -葡萄糖苷酶也得出同样的结果,诱导菌体浓度 50~g/L 发酵条件下获得的酶活显著高于60~g/L 的菌体浓度。
酵母细胞壁结构坚韧,由3层物质构成,外层为甘露聚糖(Mannan),内层为葡聚糖(Glucan),中间层为甘露聚糖蛋白,必须使用外力才能进行破除]。超声波破碎法是常用的酵母细胞破壁方法,利用超声波在液体介质中的空化作用产生的局部高温高压来破碎细胞。该方法效率高、操作简单且液量损失少,但容易导致液体局部温度过高,使目标蛋白失活[²2]。因此,毕赤酵母异源表达菌株破壁条件的优化是SbES蛋白粉剂生产的关键部分之一。本研究首先筛选了超声功率,发现功率达到 300\;W 后,破碎上清酶活达到 480\ IU/L ,而功率增加至400和 500~W 后,上清酶活不再增加。功率的增加会带来菌体悬浮液温度过高,导致目的蛋白存在失活的风险,所以 300~W 是目的菌株最适的超声破碎功率。菌体浓度也是影响超声波破碎效率的主要因素之一,通过设置不同菌体浓度发现, 15% 菌体浓度下破碎的上清酶活较高。在此基础上,进一步筛选了超声破碎时间,发现在超声破碎时间为 20\;min 时,上清酶活仅为 390\ IU/L ,增加至 40\;min 后上清酶活可提高至 530\;\:IU/L ,但增加至 50~\min 后,上清酶活不再增加,同样考虑超声处理时间过长带来的蛋白变性问题,选择 40\ min 为最适超声破碎时间。
喷雾干燥是用雾化器将蛋白液分散为液滴,利用热空气将物料迅速干燥成粉。在干燥过程中,瞬时高温会使物料发生最小程度的变性,且投资小、生产效率高,适用于工厂化生产[23]。助剂类型与干燥条件对蛋白活性的保持具有显著影响。喷雾干燥常用助剂以碳水化合物聚合物为主,包括淀粉及其衍生物、食品胶体、纤维素及其衍生物等。喷雾干燥的进风温度直接影响到物料的干燥程度,进风温度过高,会引起整个干燥塔内的温度升高,物料中水分的蒸发速率也会迅速提高,容易造成液滴膨胀、破碎或分裂,酶蛋白的空间结构也容易变化,从而引起酶活力降低,导致酶活力回收率降低;进风温度低,会导致物料因干燥程度不够、水分超标而造成粘壁,粘附于管道和干燥塔中,引起出粉率降低24。本研究通过筛选多个干燥助剂并优化助剂比例发现, 5% 的麦芽糊精对保持SbES蛋白活性效果较好。进风口温度筛选发现, 140 °C 酶活回收率较高,但出粉量低于 150 °C
本研究的不足主要在于SbES蛋白异源表达菌株 P. pastorisX-33(pRICZA::SbES)为内源性表达,将细胞破碎后才能获得目的蛋白,且酵母菌株细胞壁坚韧,为后续高密度发酵条件优化、粉剂制备工艺研发等实验增加了困难,严重影响了实验进展。今后将在本研究基础上,综合优化密码子、转录因子和翻译相关原件,进一步提高SbES蛋白表达量,并筛选适合的外泌信号肽,构建外泌表达的毕赤酵母异源表达菌株,使该激发子蛋白的表达、纯化和粉剂加工更加便利,最终实现SbES激发子蛋白的商品化开发。
参考文献
[1]Montesano M,Brader G, Tapio Palva E.Pathogen derivedelicitors: Searching for receptors in plants[J]. MolecularPlant Pathology, 2003, 4(1): 73-79.
[2]汪惠,谢琳琳,刘文波,等.2种harpin内源表达对短短芽孢杆菌HAB-5菌株抑制和促生能力的影响[J].江苏农业科学,2015,43(11):161-166.
[3]盛世英,周强,邱德文,等.植物免疫蛋白制剂阿泰灵诱导小麦抗病增产效果及作用机制[川].中国生物防治学报,2017, 33(2): 213-218.
[4]李泽,梁颖博,段佳琪,等.蛋白激发子PevD1诱导本生烟植保素辣椒醇的产生和积累[J].植物保护,2021,47(1):31-39.
[5]齐悦,吕峻元,张悦,等.小麦叶锈菌效应蛋白Pt18906激发 TcLr^{27+}31 的双层防御反应[].中国农业科学,2020,53(12): 2 371-2 384.
[6]杨晨宇,梁颖博,贾慧慧,等.灰葡萄孢蛋白激发子BcGs1诱导番茄抗病性及功能结构域鉴定[川.植物病理学报,2017,47(5):612-618.
[7]Hael-Conrad V,Perato S M,Arias M E, et al.The elicitorprotein AsES induces a systemic acquired resistance re-sponse accompanied by systemic microbursts and mi-cro-hypersensitive responses in Fragaria ananassa[J]. Mo-lecular Plant-Microbe Interactions, 2018, 31(1): 46-60.
[8]杨扬,陈奕鹏,蔡吉苗,等.帚枝霉属内生真菌激发子蛋白 SbES诱导辣椒抗棒孢叶斑病作用机理[J.植物保护学报,2023,50(5):1336-1346.
[9]李超萍,杨扬,时涛,等.来源于帚枝霉属真菌的激发子蛋白在防治木薯细菌性菱病中的应用:CN111713511B[P/OL]. 2021-08-31, http://epub.cnipa.gov.cn/.
[10]Chalfoun N R, Durman S B, Budeguer F, et al. Developmentof PSP1, a biostimulant based on the elicitor AsES for diseasemanagement in monocot and dicot crops[J]. Frontiers inPlant Science, 2018, 9: 844.
[11]邱德文,杨秀芬.蛋白质农药研究与产业化进展[].中国农业科技导报,2006,8(6):1-4.
[12]张动军,王香芝,张付强,等.新型植物免疫激活蛋白维大力(VDAL)对小麦抗逆及产量的影响[J].农业科技通讯,2020(10): 86-89.
[13]薛建光,王琳,张付强,等.新型植物免疫激活蛋白维大力在马铃薯上的增产抗病作用初探[小.农业与技术,2020,40(24): 4-7.
[14]袁京京.植物激活蛋白工厂化生产及田间应用研究[D]北京:中国农业科学院,2009.
[15]刘权,李广悦,曾洪梅,等.蛋白激发子PeaT1的高密度发酵及生理功能检测[J].生物技术通报,2009,25(8):162-165.
[16]张强,刘祥臣,余贵龙,等.不同浓度阿泰灵对再生稻两优 6326秧苗素质和纹枯病抗性及产量的影响[J.江苏农业科学,2019,47(15):130-133.
[17]郝建宇,王伟军,陈文朝,等.生物药剂阿泰灵结合化学农药减施在^脆光'葡萄中的应用[J].中国果树,2018(4):66-68.
[18]茹扎·也里扎提,杨宇.毕赤酵母中外源蛋白表达量的提升策略[J].生物技术通报,2024,40(3):118-134.
[19]继清,曹丁,张宜靖,等.巴斯德毕赤酵母表达人溶菌酶分批发酵工艺研究[J].轻工科技,2019,35(8):24-26.
[20]汪飞,吴敬,陈晟.重组毕赤酵母生产 β^{*} 葡萄糖苷酶发酵条件优化及固定化研究[].食品科学技术学报,2019,37(3): 41-47.
[21]石豪磊,赵国正,孔祥君,等.碱法破壁-酶法提取葡萄酒废酵母细胞壁多糖的工艺研究[J].中国酿造,2018,37(12):82-86.
[22]杨翠竹,李艳,阮南,等.酵母细胞破壁技术研究与应用进展[J].食品科技,2006,31(7):138-142.
[23]鲁玉佳,田路,胡亚平,等.喷雾干燥法制备双孢菇蛋白粉工艺优化及其营养价值评价[].食品工业科技,2018,39(10): 90-94+104.
[24] Verma A, Singh S V. Spray drying of fruit and vegetablejuices—a review[J]. Critical Reviews in Food Science andNutrition, 2015, 55(5): 701-719.
基于 MAGIC群体的烟草抗黑胫病全基因组关联分析
潘怡1.2 刘洪泰”吉白璐1,2 王红²黄立钰’程立锐2(1.云南大学农学院云南昆明650504;2.中国农业科学院烟草研究所山东青岛266101;3.云南省保山市烟草公司腾冲分公司
云南保山679100)
摘要由烟草疫霉引起的黑胫病是烟草主要病害之一,严重影响烟叶的产量和品质。为了进一步明确烟草黑胫病抗性遗传基础,发掘烟草黑胫病抗性位点,采用8亲本组配的 MAGIC(Multiparent AdvancedGeneration Inter-Cross)群体为试验材料,在苗期和大田期调查亲本及后代群体的黑胫病抗性;利用 430~~K~ 烟草SNP(SingleNucleotidePoly-morphism)芯片鉴定基因型,并对烟草黑胫病不同生育期抗性进行全基因组关联分析(Genome-WideAssociation Study,GWAS)。结果显示,在苗期和大田期重复定位到2个与黑胫病病情指数及发病率相关的黑胫病抗性主效QTL(QuantitativeTraitLocus),分别位于7号和14号染色体,最显著SNP物理位置分别在7号染色体175557310bp和14号染色体7074846bp;其中,7号染色体定位结果与前人研究结果一致,14号染色体定位结果为烟草黑胫病抗性新位点。本研究为深人挖掘烟草抗黑胫病主效基因和开展抗性分子标记辅助改良提供参考。
关键词MAGIC群体;烟草;黑胫病;GWAS
35.72 文献标识码A DOI:10.12008/j.issn.1009-2196.20:
Genome-Wide Association Study for Black Shank Disease Resistance in Tobacco Based on the MAGIC Population
PAN Yi 12 LIU Hongtai? JI Bailul2 WANG Hong² HUANG Liyu' CHENG Lirui2 (1. School of Agriculture, Yunnan University, Yunnan, Kunming 650504, China; 2. Tobacco Research Institute, Chinese Academy of Agricultural Sciences, Qingdao, Shandong 266101, China; 3. Yunnan Tobacco Company Baoshan Company Tengchong Branch, Yunnan,Baoshan679100,China)
Abstract Black shank, one of the paramount diseases affecting the yield and quality of tobacco, is caused by Phytophthora nicotianae.To further clarify the genetic basis of black shank disease resistance and discover new resistance loci for this disease, a MAGIC (Multiparent Advanced Generation Inter-Cross) population with eight parents was used as the material, and the resistance of the parental and offspring populations'resistance to black shank disease was investigated at the seedling and field stages. We identified the genotypes of the MAGIC population via a 430~K tobacco SNP (Single-Nucleotide Polymorphism) chip and conducted a genome-wide association study (GWAS)for tobacco black shank resistance. The results revealed that two major QTLs (Quantitative Trait Locus) related to the disease index and incidence rate of black shank resistance, which are located on chromosomes 7 and 14, were repeatedly identified at the seedling stage and field stage. The most significant SNPs were physically located on chromosome 7 at 175 557 310 bp and chromosome 14 at 7 074 846 bp, respectively. The localization of chromosome 7 is in line with the results of previous studies, and the localization of chromosome 14 is a new locus for tobacco black shank disease resistance. The results of this study provide reference for further exploration of major genes conferring resistance to tobacco black shank disease and the implementation of resistance molecular marker-assisted improvement.
Keywords MAGIC population; tobacco; black shank; GWAS
烟草黑胫病是由烟草寄生疫霉菌(Phytophthoraparasitica var.nicotianae)引发的土传病害[-2],在整个烟草生长发育期均可发生,以苗期和大田期危害最为严重,每年给国家和烟农带来巨大经济损失[3-4]。迄今为止,烟草黑胫病防治策略主要包括抗性品种培育与利用、轮作种植和药剂防治等。其中,培育抗病品种是防治黑胫病最有效的途径之一。
烟草黑胫病抗源主要有4个,分别为雪茄烟品种Florida301和Beinhart1000、野生种蓝茉莉叶烟草(N.plumbaginifolia)和长花烟草(N.longiflora)。烟草黑胫病抗性育种以利用水平抗源为主,其抗性遗传属于数量遗传,由微效多基因控制[6-8]。目前,国内外有关烟草抗黑胫病的QTL定位研究有不同程度的进展。Vontimitta等[9利用抗性品种Beinhart1000与感病品种Hicks杂交,构建了双单倍体加倍群体,结果显示,与黑胫病抗性相关的QTL被定位在6个不同位置,并以位于4号连锁群和8号连锁群上的QTL 贡献率最高。Xiao 等[10]以抗病材料Florida301和感病材料Hicks为亲本构建重组自交系,定位到11个与黑胫病抗性相关的QTL。郭璇[8利用抗病品种Beinhart1000-1分别与2个感病品种小黄金1025、SamsunNN配置杂交组合,定位到27个黑胫病抗性相关QTL,位于 12 号连锁群上的 QTL 效应值最大。Zhang等[同样以 Beinhart1000-1 和小黄金1025 为亲本,基于所构建的不同群体材料在温室和田间环境下重复定位到qBS7和qBS17两个抗黑胫病主效QTL。Gong 等2]以高抗材料云烟85与易感材料大白金599杂交配置了177份 \operatorname{F}_{7:8-9} 重组自交系制图群体,在多环境下重复定位到gBS7和qBS14两个与黑胫病抗性相关的QTL,分别解释了16.48%~62.20% 和 3.94%~11.29% 的表型变异。到目前为止,虽然关于烟草黑胫病抗性的主效位点定位已经有所报道,但是大多研究采用的是基于双亲杂交构建的遗传群体,群体遗传多样性和重组率较低,进而直接影响了定位结果的准确性和进一步的育种利用。
多亲本高世代互交群体(MAGIC群体)是近年发展起来的新一代遗传作图及育种群体,最初是以研究动物复杂性状遗传为目标而构建的基于多亲本的重组近交群体,随后其构建方法被衍生到植物中应用3]。在遗传研究上,MAGIC群体具有显著优势,其内包含的大量可利用的多样性遗传变异可同时定位多个QTL和主效基因,既保证了群体定位的精度[14],又减少了群体结构和稀有等位变异等方面的问题,切实做到了育种群体和定位群体的有机整合[3]。本研究以8亲本组配的MAGIC 群体为材料,分别在苗期和大田期鉴定黑胫病抗性,结合重测序技术鉴定得到的基因型数据,对烟草黑胫病抗性进行GWAS分析,发掘稳定表达的显著SNP位点,为进一步挖掘烟草抗黑胫病主效基因和开展品种抗性改良提供参考。
1 材料与方法
1.1 材料
以VAM(白肋烟)、Beinhart1000-1(雪茄烟)、Florida301(雪茄烟)、Samsun(香料烟)、小花青(晾晒烟)、塘蓬(晾晒烟)、红花大金元(烤烟)、Basma(香料烟)等8种不同烟草类型种质(均由中国烟草种质资源库提供)为亲本配置不完全双列杂交,依次进行两亲本 F_{1} 相互配组、四亲本 F_{l} 相互配组,将所获得的含有八亲本遗传背景的杂交组合 F_{1} 种子等量混合,种植后自交收获 F_{2} ,之后采用单粒传法自交4代,获得8亲本烟草MAGIC群体,包含600个株系(图1)。本研究所鉴定的苗期材料共利用其中的526个,大田期材料共利用其中的496个,群体材料均种植于中国农业科学院烟草研究所青岛试验基地。

图1MAGIC群体构建示意图
试验菌种为烟草黑胫病菌0号生理小种,由中国农业科学院烟草研究所遗传育种科室提供。
1.2方法
1.2.1试验设计苗期供试材料种植于试验基地温室大棚,待播种后幼苗长至3\~4片真叶时,移栽于假植盘中。每份材料设置2个重复,每个重复25株。采用刘洪泰5]方法制备菌谷。当供试材料长至4叶1心期,通过创伤烟草植株茎基部接种菌谷,统一控制好用量,接种7d后调查发病情况。
大田期供试材料种植于试验基地黑胫病病圃,田间种植行距和株距分别为60、 40\ {cm} ,每行10棵,设2次重复。当烟株长至旺长期,通过创伤烟株茎基部统一接种菌谷,接种30d后调查发病情况。
1.2.2黑胫病发病调查烟草黑胫病调查方法和分级严格按照国家标准(GB/T23222一2008)开展。以株为单位进行的病情分级鉴定,根据受害程度划分为0、1、3、5、7和9级[6];以群体为单位进行抗性等级鉴定,根据病情指数(简称病指)划分为抗病(病指25以下)、中抗(病指25.1~50.0 )、中感(病指50.1\~75.0)和感病(病指75以上)[7。病情指数和发病率计算公式如下。
>(病级级代表 x 该级烟草株数)病情指数 = x100 调查总株数 x 最高级高级发病率 = 染病株数 ×100%调查总株数
1.2.3MAGIC群体构建及基因型鉴定针对MAGIC群体的8亲本及群体内各材料,摘取株系内烟草单株幼嫩叶片进行混合取样,采集时最大程度保持样本一致性。采用SLS法[5提取不同样品基因组DNA,确保达到芯片检测要求。烟草全基因组SNP分型检测基于 430 K 烟草SNP芯片进行[18],具体参照张剑锋等1"流程。构建图谱时排除质量较差或数据缺失超过 10% 的SNP标记,共计鉴定到93766个有效SNP适用于群体材料基因型分析。
1.2.4性状与标记的全基因组关联分析基于苗期和大田期质控后的基因型数据及抗黑胫病表型数据,利用TASSEL软件混合线性模型进行GWAS[18],以 -log10P>=slant5 为标准筛选显著相关的SNP并绘制相应的Manhattan-Plot。
2 结果与分析
2.1表型变异及SNP标记多态性
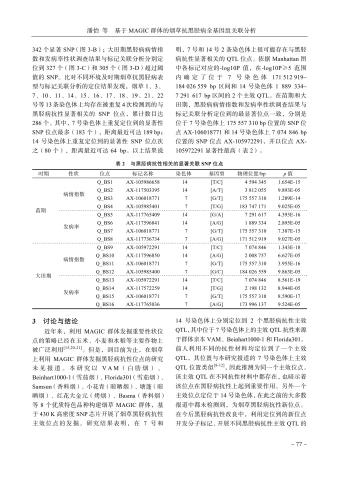
在苗期和大田期,亲本及MAGIC群体内各株系材料的黑胫病抗性有所差异(图2)。亲本Beinhart1000-1对烟草黑胫病的抗性最好,苗期病指和发病率分别为0.44和 4.00% ,大田期病指和发病率分别为1.78和 16.00% ;抗性较好是Florida301。VAM、Samsun和塘蓬在苗期和大田期黑胫病抗性总体表现为中抗,在苗期,病指变异范围为43.93\~46.90,发病率变异范围为 60.87%~ 83.33% ;在大田期,病指变异范围为20.44\~49.54,发病率变异范围为 48.00%~96.00% 。小花青的黑胫病抗性在苗期表现为中抗,而大田期表现为感病。红花大金元和Basma两个亲本的黑胫病抗性在苗期表现为中感,病指和发病率变异范围分别为66.67\~67.29、 76.00%~93.75% ;大田期表现为中抗,病指和发病率变异范围分别为29.33\~31.11、64.00°/_{0}~88.00°/_{0} 。后代群体各株系苗期病指平均值为26.83、变异系数为 80.57% ,发病率平均值为 47.52% 、变异系数为 50.46% ,总体呈现连续变异趋势;相类似的,大田期各株系病指平均值为20.71、变异系数为 101.35% ,发病率平均值为43.46% 、变异系数为 61.57% 。总体而言,亲本及MAGIC群体间苗期和大田期黑胫病抗性存在显著性差异,呈现数量性状遗传分布特征。
2.2黑胫病抗性与SNP的全基因组关联分析(GWAS)
利用 430~K 烟草SNP芯片鉴定群体材料基因型,共获得93766个较为均匀分布在烟草24条染色体上的高质量多态性SNP标记,可覆盖烟草全基因组长度 4 019.64~Mb ,平均SNP标记间的物理距离为 42.86~Kb 。其中,22号染色体上的标记最多(7316个),15号染色体上的标记最少(2301个)(表1)。
基于TASSEL软件混合线性模型,对苗期和大田期质控后的基因型数据及抗黑胫病表型数据进行全基因组关联分析,以 {*}\log10P>=5 为标准筛选显著相关的SNP。结果(图3)显示,苗期黑胫病病情指数与标记关联分析定位到336个显著SNP(图3-A),发病率与标记关联分析定位到

MAGIC群体苗期共覆盖526份试验材料,大田期共覆盖496份试验材料。A.苗期病情指数;B.苗期发病率;C.大田期病情指数;
D.大田期发病率。MAGIC群体所含8个亲本:P1(VAM);P2(小花青);P3(Beinhartl000-1);P4(Basma);P5(Samsun);P6(Florida301);P7(塘蓬);P8(红花大金元)。
| 染色体 | 标记数目 | 起始位置/Mb | 终止位置/Mb | 覆盖染色体距离/Mb | 平均标记间隔/Kb |
| 1 | 3068 | 0.08 | 227.35 | 227.26 | 74.08 |
| 2 | 4 446 | 0.53 | 136.67 | 136.14 | 30.62 |
| 3 | 3309 | 0.54 | 217.78 | 217.24 | 65.65 |
| 4 | 4 779 | 0.01 | 158.19 | 158.18 | 33.10 |
| 5 | 2 641 | 1.84 | 169.47 | 167.63 | 63.47 |
| 6 | 5 712 | 0.92 | 221.75 | 220.83 | 38.66 |
| 7 | 4 004 | 0.11 | 187.98 | 187.88 | 46.92 |
| 8 | 3522 | 0.74 | 212.90 | 212.16 | 60.24 |
| 9 | 4325 | 2.98 | 133.31 | 130.33 | 30.13 |
| 10 | 4 069 | 0.09 | 170.24 | 170.16 | 41.82 |
| 11 | 3166 | 0.72 | 182.34 | 181.62 | 57.37 |
| 12 | 2 914 | 0.04 | 137.06 | 137.02 | 47.02 |
| 13 | 2 972 | 0.01 | 141.13 | 141.12 | 47.48 |
| 14 | 6 412 | 0.01 | 123.90 | 123.89 | 19.32 |
| 15 | 2301 | 0.01 | 141.42 | 141.41 | 61.46 |
| 16 | 3791 | 0.01 | 178.60 | 178.59 | 47.11 |
| 17 | 2 749 | 0.05 | 163.01 | 162.96 | 59.28 |
| 18 | 2722 | 2.03 | 166.79 | 164.76 | 60.53 |
| 19 | 3498 | 0.22 | 188.79 | 188.57 | 53.91 |
| 染色体 | 标记数目 | 起始位置/Mb | 终止位置/Mb | 覆盖染色体距离/Mb | 平均标记间隔/Kb |
| 20 | 5 229 | 0.47 | 168.38 | 167.92 | 32.11 |
| 21 | 3930 | 0.09 | 102.04 | 101.95 | 25.94 |
| 22 | 7316 | 0.17 | 243.47 | 243.30 | 33.26 |
| 23 | 4 090 | 9.24 | 153.13 | 143.89 | 35.18 |
| 24 | 2801 | 0.04 | 114.85 | 114.82 | 40.99 |
| 合计 93766 | 4 019.64 | 42.86 | |||
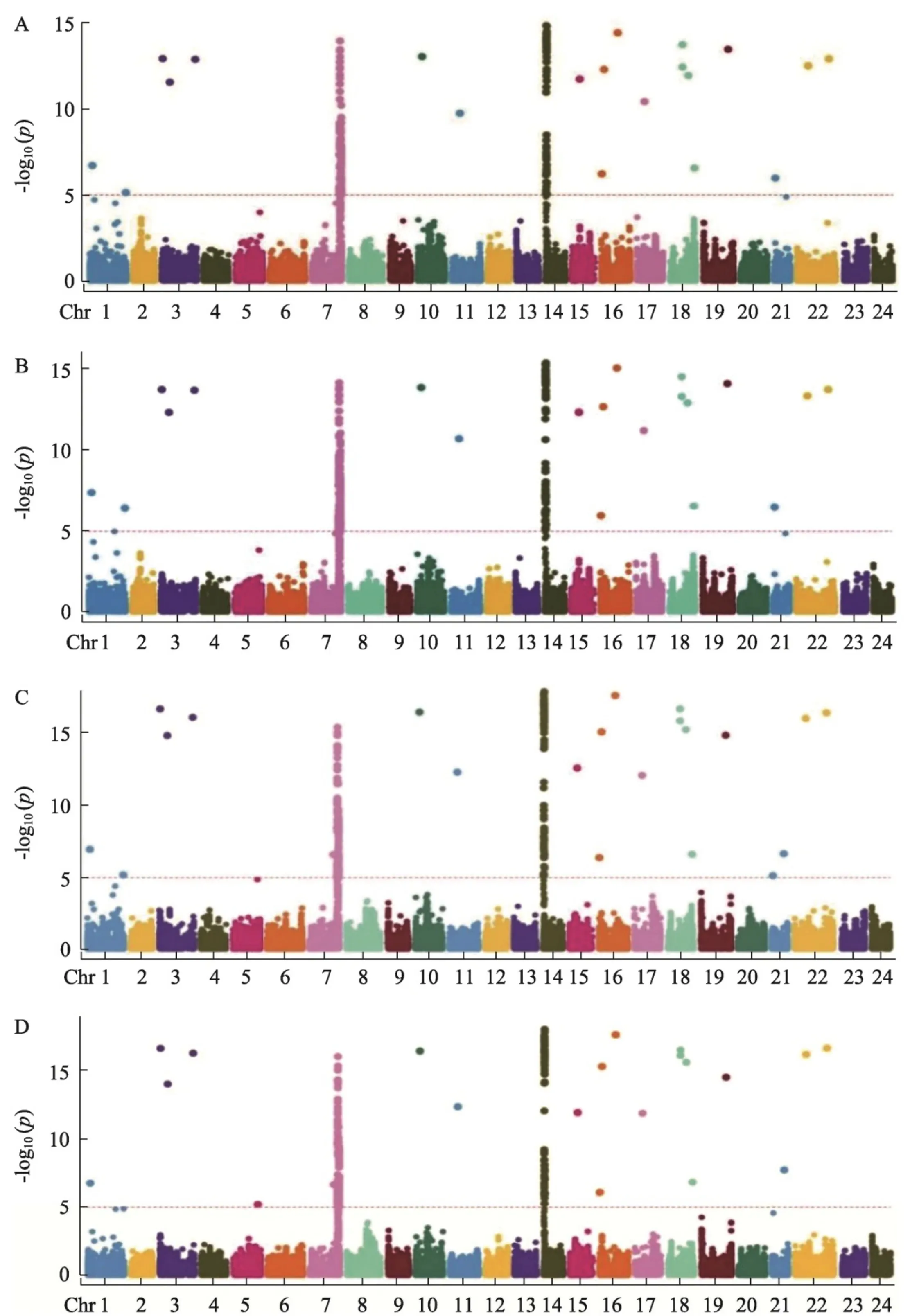
将苗期526份试验材料和大田期496份试验材料抗黑胫病表型与93766个SNP标记进行全基因组关联分析。A.苗期病情指数与标记关联分析曼哈顿图;B.苗期发病率与标记关联分析曼哈顿图;C.大田期病情指数与标记关联分析曼哈顿图;D.大田期发病率与标记关联分析曼哈顿图;阈值设置为5。
342个显著SNP(图3-B);大田期黑胫病病情指数和发病率性状调查结果与标记关联分析分别定位到327个(图3-C)和305个(图3-D)超过闽值的SNP。比对不同环境及时期烟草抗黑胫病表型与标记关联分析的定位结果发现,烟草1、3、7、10、11、14、15、16、17、18、19、21、22号等13条染色体上均存在被重复4次检测到的与黑胫病抗性显著相关的SNP位点,累计数目达286个。其中,7号染色体上重复定位到的显著性SNP位点最多(183个),距离最近可达 189 {\mathsf{b p}} 14号染色体上重复定位到的显著性SNP位点次之(80个),距离最近可达 64~{\mathfrak{b}}{\mathfrak{p}} 。以上结果说明,7号和14号2条染色体上很可能存在与黑胫病抗性显著相关的QTL位点。依据Manhattan图中各标记对应的-log10P值,在 -\log10\mathbb{P}>=slant5 范围内确定了位于7号染色体 171\;512\;919~ 184026559bp区间和14号染色体 1\;\;889\;\;334~ 7291617bp区间的2个主效QTL。在苗期和大田期,黑胫病病情指数和发病率性状调查结果与标记关联分析定位到的最显著位点一致,分别是位于7号染色体上175557310bp位置的SNP位点AX-106018771和14号染色体上7074846bp位置的SNP位点AX-105972291,并以位点AX-105972291显著性最高(表2)。
| 时期 | 性状 | 位点 | 标记名称 | 染色体 | 基因型 | 物理位置/bp | P值 |
| 苗期 | 病情指数 | Q_BS1 | AX-105986658 | 14 | [T/C] | 4594345 | 1.654E-15 |
| Q_BS2 | AX-117503395 | 14 | [A/T] | 3812055 | 9.893E-05 | ||
| Q_BS3 | AX-106018771 | 7 | [G/T] | 175557310 | 1.289E-14 | ||
| Q_BS4 | AX-105985401 | 7 | [T/G] | 183 747 171 | 9.025E-05 | ||
| Q_BS5 | AX-117765409 | 14 | [G/A] | 7291 617 | 4.393E-16 | ||
| Q_BS6 | AX-117596841 | 14 | [A/G] | 1889334 | 2.895E-05 | ||
| 发病率 | Q_BS7 | AX-106018771 | 7 | [G/T] | 175 557 310 | 7.387E-15 | |
| Q_BS8 | AX-117736734 | 7 | [A/G] | 171512919 | 9.027E-05 | ||
| Q_BS9 | AX-105972291 | 14 | [T/C] | 7.074846 | 1.343E-18 | ||
| 大田期 | 病情指数 | Q_BS10 | AX-117596850 | 14 | [A/G] | 2008757 | 6.627E-05 |
| Q_BS11 | AX-106018771 | 7 | [G/T] | 175557310 | 3.955E-16 | ||
| Q_BS12 | AX-105985400 | 7 | [G/C] | 184 026559 | 9.863E-05 | ||
| 发病率 | Q_BS13 | AX-105972291 | 14 | [T/C] | 7074846 | 8.561E-19 | |
| Q_BS14 | AX-117572259 | 14 | [T/G] | 2 198 132 | 8.944E-05 | ||
| Q_BS15 | AX-106018771 | 7 | [G/T] | 175557310 | 8.590E-17 | ||
| Q_BS16 | AX-117765036 | 7 | [A/G] | 173 996 137 | 9.524E-05 |
3 讨论与结论
近年来,利用MAGIC群体发掘重要性状位点的策略已经在玉米、小麦和水稻等主要作物上被广泛利用[5,20-2]。但是,到目前为止,在烟草上利用MAGIC群体发掘黑胫病抗性位点的研究未见报道。本研究以VAM(白肋烟)、Beinhart1000-1(雪茄烟)、Florida301(雪茄烟)、Samsun(香料烟)、小花青(晾晒烟)、塘蓬(晾晒烟)、红花大金元(烤烟)、Basma(香料烟)等8个优质特色品种构建烟草MAGIC群体,基于 430~K 高密度SNP芯片开展了烟草黑胫病抗性主效位点的发掘。研究结果表明,在7号和
14号染色体上分别定位到2个黑胫病抗性主效QTL,其中位于7号染色体上的主效QTL抗性来源于群体亲本VAM、Beinhart1000-1和Florida301。前人利用不同的抗性材料均定位到了一个主效QTL,其位置与本研究报道的7号染色体上主效QTL位置类似I8-12],因此推测为同一个主效位点。该主效QTL在不同抗性材料中都存在,也暗示着该位点在黑腔病抗性上起到重要作用。另外一个主效位点定位于14号染色体,在此之前的大多数报道中都未检测到,为烟草黑胫病抗性新位点。在今后黑胫病抗性改良中,利用定位到的新位点开发分子标记、开展不同黑胫病抗性主效QTL的分子聚合改良是一种有效的育种策略。此外,本研究分别在苗期和大田期开展了烟草黑胫病抗性遗传分析和定位,结果表明,位于7号和14号染色体上的2个主效QTL在苗期和大田期均能被稳定检测到,说明在苗期和大田期2个时期烟草黑胫病抗性遗传机制类似。后续研究将进一步利用所筛选到的与黑胫病抗性显著相关的SNP位点进行区间精细定位、候选基因挖掘和功能解析,同时,利用开发的分子标记,开展非等位抗性基因的聚合研究,推进烟草抗黑胫病品种改良。
作为典型的异源四倍体,烟草基因组大小约4.5Gb,其中重复序列含量可达基因组序列的 60%~ 80%^{[22]} ,这无疑给烟草基因组的测序和拼接带来巨大挑战[23]。本研究中,在烟草1、3、7、10、11、14、15、16、17、18、19、21、22号等13条染色体上均多次重复检测到与黑胫病抗性显著相关的SNP位点,但仅确认了位于7号和14号染色体上的2个主效QTL,究其原因是黑胫病抗性遗传机制复杂,同时受主效基因和微效多基因控制。与此同时,GWAS分析基于二代测序技术,还受DNA序列读长及参考基因组拼接错误等因素的影响。目前,不同拼接版本的基因组数据库在序列长度、重复类型数量及功能基因定位等方面亦存在较大差异,这严重影响了烟草遗传分析和基因功能解析[24-25]。随着基因组测序技术的不断发展,复杂基因组T2T级别的基因组组装已成为可能。因此,在后续烟草重要性状遗传定位及基因功能解析过程中,需要优先考虑完善目前烟草参考基因组质量。
参考文献
[1] 赵辉,王喜英,刘国权,等.烟草黑胫病发生因素及综合防治研究进展[J].湖南农业科学,2020(11):99-103.
[2] 马国胜,高智谋.烟草黑胫病菌培养性状的研究[川.中国农业科学,2007,40(3):512-517.
[3] 王文静,王晓强,许永幸,等.烟草黑胫病菌分子生物学研究进展[J].中国烟草科学,2021,42(3):90-94.
[4] 赵亚南,黄大野,杨丹,等.烟草黑胫病研究进展[J].湖北农业科学,2022,61(S1): 25{-}28{+}66.
[5] 金子纯,王爱国,魏珂,等.烟草黑胫病的发病规律及综合防治技术[J].现代农业科技,2017(17):122-123.
[6] 戴彬.烟草黑胫病抗性遗传规律分析及其SSR分子标记的筛选[D].长沙:湖南农业大学,2018.
[7]王佩,张强,孙渭,等.烟草黑胫病抗性基因分子检测及其抗性特性分析[J].分子植物育种,2019,17(20):6743-6 750.
[8]郭璇.烟草黑胫病抗性遗传分析及主效位点发掘[D].青岛:青岛农业大学,2017.
[9]Vontimitta V, Lewis R S. Growth chamber evaluation of atobacco “beinha rt 1000°x “Hicks’ mapping population forquantitative trait loci affecting resistance to multiple races ofPhytophthora nicotianae[]. Crop Science, 2012,52(1): 91-98.
[10] Xiao B, Drake K, Vontimitta V, et al. Location of genomicregions contributing to Phytophthora nicotianae resistance intobacco cultivar Florida 301[J]. Crop Science, 2013, 53(2):473-481.
[11] Zhang Y S, Guo X, Yan X X, et al. Identification of stablyexpressed QTL for resistance to black shank disease in to-bacco (Nicotiana tabacum L.) line Beinhart 1000-1[J]. TheCrop Journal, 2018, 6(3): 282-290.
[12] Gong D P, Chen M L, Sun Y, et al. Fine mapping of QTLsfor resistance to Phytophthora nicotianae in flue-cured to-bacco using a high-density genetic map[J]. Molecular Breed-ing, 2020, 40(5): 45.
[13]荣克伟,柳波娟,卢跃磊,等.MAGIC 群体的遗传特征及其在作物耐逆研究上的应用[].作物杂志,2022(3):9-19.
[14]汪夏超,栾悦,李春林,等.不同遗传群体在数量性状基因定位分析中的应用概述[].蚕学通讯,2023,43(2):14-24.
[15]刘洪泰.基于 MAGIC 群体的烟草黑胫病抗性遗传分析[D].北京:中国农业科学院,2020.
[16]蒋彩虹,王元英,高亭亭,等.雪茄烟Beinhart1000-1抗赤星病和黑胫病基因的QTL定位[C]//CORESTA2014年大会入选论文集,2014-12.中国北京:[出版社不详],2014:156-164.
[17]苗圃,王海涛,李淑君,等.河南省烟草黑胫病菌生理小种鉴定[J].西北农业学报,2013,22(10):204-207.
[18]孙滢,姜自鹏,刘洪泰,等.烟草开花期全基因组关联分析[].中国烟草科学,2020,41(6):1-6.
[19]张剑锋,罗朝鹏,何声宝,等.应用SNP标记分析24份烟草品种的遗传多样性[J].烟草科技,2017,50(11):1-8.
[20] Ravelombola W, Shi A, Huynh B L, et al. Genetic architec-ture of salt tolerance in a Multi-Parent Advanced GenerationInter-Cross (MAGIC) cowpea population[]. BMC Ge-nomics, 2022, 23(1): 1-7.
[21]钟丽萍,王尖,吴晓花,等.基于MAGIC群体瓜白粉病抗性的全基因组关联分析[].浙江农业学报,2023,35(10):2 398-2 407.
[22]姜自鹏,赵会纳,苑广迪,等.烟草株高和叶数性状QTL定位及候选基因预测[].中国烟草科学,2022,43(2):1-6.
[23]王元英.烟草基因组知识篇:1.基因组与烟草基因组计划[J.中国烟草科学,2010,31(1):81-82.
[24]张玉婵,田嘉树,王晓丽,等.基于文献计量学的烟草基因组研究知识图谱分析[J.中国烟草学报,2022,28(4):85-95.
[25]卢鹏,金静静,李泽锋,等.基于第三代测序技术的基因组组装方法及其在烟草中的应用[川.烟草科技,2018,51(2): 87-94.
(责任编辑林海妹)
基于机器学习算法的森林病虫害遥感模型对比研究
郑绍鑫」何江」封成」陈积标’潘兴建’李军集2.3[1.广西壮族自治区国有六万林场广西玉林537000;2.广西壮族自治区林业科学研究院/广西特色经济林培育与利用重点实验室/广西木本香料工程技术研究中心广西南宁530002;3.中国地质大学(北京)信息工程学院北京1000831
摘要森林病虫害严重威胁森林的生态功能与经济效益,如何有效识别森林病虫害并对其危害程度进行估测,对维护森林资源的健康和可持续利用至关重要。遥感技术能够实时、大范围连续动态获取植被对环境胁迫的光谱响应信息,广泛应用于森林病虫害监测。以广西六万林场森林公园为实验区,将地面调查数据及2022一2023年Sentinel-2遥感影像作为基础数据源,选取对植被颜色变化敏感的植被衰减指数(PSRI)、对植被结构变化敏感的结构不敏感色素指数(SIPI)、归一化差值红边指数(NDRE)及对植被功能变化敏感的植被光合有效辐射吸收系数(FAPAR)、归一化差值水体指数(NDWI)作为森林病虫害监测的光谱特征参数,分别利用支持向量机(SVM)和随机森林(RF)两类机器学习算法建立森林病虫害胁迫遥感监测模型,从总体精度、Kappa系数等指标对模型精度进行评价。结果表明:(1)广西六万林场森林公园的病虫害具有集聚分布、连片蔓延特点,多发生于11一12月林场森林公园北部与中部地区;(2)利用RF与SVM进行森林病虫害遥感分类的总体精度分别为 63.51%~92.21% 52.70%~81.82% ,Kappa系数分别为 0.54~ 0.90、0.41\~0.77;(3)相比SVM算法,基于RF的森林病虫害遥感监测模型精度更高,更有利于森林病虫害的监测识别。研究结果为探讨实验区的森林病虫害时空分布提供基础数据,为森林病虫害防治提供科学依据,同时为大尺度森林病虫害遥感监测提供参考。
关键词森林病虫害;Sentinel-2影像;敏感光谱参数;随机森林;支持向量机
Comparative Research on Remote Sensing Models for Forest Pests and Diseases via Machine Learning Algorithms
ZHENG Shaoxin' HE Jiang' FENG Cheng' CHEN Jibiao' PAN Xingjian' LI Junji2.3
[1. Guangxi State-owned Liuwan Forest Farm, Yulin, Guangxi 537000, China; 2. Guangxi Forestry Research Institute/Guangxi Key
Laboratory of Characteristic Non-wood Forest Cultivation & Utilization/Guangxi Engineering and Technology Research Center for Woody Spices, Nanning, Guangxi 530002, China; 3. School of Information Engineering, China University of Geosciences (Beijing),Beijing100083,China]
AbstractForestpests and diseases seriouslythreaten the ecologicalfunction and economicbenefits of forests.Howtoidentify forest pests and diseases effectively and assessing their severity is very important for maintaining the health and sustainable use of forest resources. Remote sensing technology can continuously and dynamically obtain the spectral response information of vegetation to environmental stress in real-time and on a large scale, so it is widely used in forest pest and disease monitoring.In this study, the Guangxi State-owned Liuwan Forest Farm was taken as the experimental area,and the ground survey data and Sentinel-2remote sensing images from 2022 to 2023 were used as the fundamental data sources.The plant senescence reflectance index (PSRI), which is sensitive to changes in vegetation function; the structure insensitive pigment index (SIPI); and the normalized difference red edge index (NDRE), which is sensitive to vegetation structure change; the fraction of absorbed photosynthetically active radiation (FAPAR); and the normalized difference water index (NDWI), which issensitivetovegetationfunctionchangeswereselectedasthespectralfeatureparametersformonitoringforestpestsand diseases. Two kinds of machine learning algorithms, support vector machine (SVM) and random forest (RF), were used to establish a remote sensing monitoring model of forest pests and disease stress,and the accuracy of the model was evaluated in terms of the overall accuracy,the Kappa coefficient,and other indicators. The results revealed that: 1) the forest pests and diseases at Liuwan Forest Farm in Guangxi were characterized by an agglomeration distribution and contiguous spread,which occurred mainly in the northern and central areas of the forest farm from November to December; 2)the overall accuracies of the RF and SVM remote sensing classifications of forest pests and diseases were 63.51%-92.21% and 52.70%-81.82% respectively, and the Kappa coefficients were 0.54-0.90 and 0.41-0.77,respectively; 3) compared with the SVM algorithm, the RF-based forest pest and disease remote sensing monitoring model has higher accuracy and is more conducive to the monitoring and identification of forest pests and diseases. The results provide fundamental data on the spatial and temporal distributions of forest pests and diseases in the experimental area, supply a scientific basis for forest pest control, and provide a referenceforremote sensingmonitoring oflarge-scaleforest pests and diseases.
Keywords forest pests and diseases; Sentinel-2 images; sensitive spectral indices; random forest; support vector machine
森林资源承担着水源涵养、空气净化、防风固沙等重要功能,对维护生态系统的稳定性及推动社会可持续发展至关重要[-3]。然而,森林病虫害的发生严重威胁森林的生态功能和经济效益,有效监测和防控森林病虫害对于维护森林资源的健康和可持续利用具有重要意义[4-5]。传统的野外人工探查虽可获取森林病虫害情况,但具有视距受限、可达性差、费时费力等缺点,卫星遥感技术具有大范围、全覆盖及动态周期性的优势,在森林病虫害研究中得到广泛应用。Sentinel-2具有高时空分辨率特性,且包含3个“红边”波段,为植被健康状况监测提供了有利条件[7-8]。
病虫害胁迫会改变树木的外部形态和内部生理功能,阻碍其生长甚至导致其死亡"。植被指数是一种利用叶冠的光学参数提取的独特光谱信号,可用于准确监测植物的光合活性、生化胁迫的变化以及其他植被压力"。为了有效监测森林病虫害,许多学者针对森林的光谱特征变化开展研究,如 Xu 等]利用叶面积指数(LAI)、叶面积指数标准误差(SEL)、归一化植被指数(NDVI)等作为光谱特征对森林病虫害进行识别;郭仲伟等2探讨叶面积指数(LAI)、归一化植被指数(NDVI)与增强型植被指数(EVI)与不同程度森林病虫害的关系;Zhang等[3]采用归一化植被指数(NDVI)和归一化差异红边指数(NDRE)表征植被叶绿素含量,以归一化水分指数(NDMI)及归一化燃烧比率(NBR)表征植被含水量,以此来预测松材线虫的易感性和风险水平;Lei等[4选取归一化植被差指数(NDVI)、优化土壤调整植被指数(OSAVI)、叶片叶绿素指数(LCI)、绿色归一化植被差指数(GNDVI)归一化差分红边指数(NDRE)5种植被指数建立模型,以确定槟榔黄叶病的严重程度。但目前研究多依据受胁迫植被形态(如LAI)、颜色(如叶绿素)变化信息,较少综合蒸腾作用、光合作用、呼吸作用等生理功能特征变化来揭示森林病虫害胁迫情况。综合参考植被的颜色、结构与功能等信息,选择合适的特征参数进行森林病虫害监测识别,可更全面地了解森林健康情况,为病虫害预防和控制提供科学依据[15-16]
除光谱特征参数外,分类方法的选择也对森林病虫害的监测效果产生影响[7。支持向量机(SupportVectorMachine,SVM)与随机森林(RandomForest,RF)算法是常用的机器学习分类算法,可对病虫害进行分类监测[8]。黄芳芳等[19]以随机森林和决策树方法,结合7个原始光谱波段对马尾松松材线虫病进行研究,平均精度在76% 以上。Zhan等[2比较了决策树、支持向量机与随机森林算法对高分影像中森林虫害的分类效果。 Xu 等2"基于无人机获取的多光谱图像提取敏感特征,并分别以支持向量机、随机森林和极端梯度增强树3类机器学习算法识别食叶害虫对毛竹林的危害。以上研究表明,机器学习法对植被病虫害监测的有效性。本研究基于Sentinel-2 影像数据,选取综合森林植被颜色、结构、功能的植被指数,构建基于支持向量机及随机森林算法的森林病虫害遥感监测模型,并分析对比2种方法的结果,为森林病虫害的快速、大面积监测提供技术支持,为深入开展病虫害遥感监测应用奠定基础。
1 材料与方法
1.1材料
1.1.1研究区概况研究区位于广西壮族自治区玉林市福绵区的广西国有六万林场森林公园(以下简称森林公园),地处玉林市福绵区、博白县、兴业县和钦州市浦北县交界处,如图1所示。其属亚热带季风气候区,季节性变化显著,气候暖热、气温较高、热量充足、雨量充沛,年平均气温约 22 °C ,年平均降水量约 1\ 500\mm
截至2022年,森林公园现有经营面积 5.93x
10^{4} hm^{2} ,主要种植八角,另有少量按树、松树等分布,森林覆盖率达 93% 。近年来,受气候环境、种植条件等的影响,公园内病虫害发生频繁,其中八角炭疽病、尺等病虫害危害严重"",对森林的健康生长产生巨大影响,制约着园区林业产业的持续健康发展。
1.2方法
1.2.1数据收集及预处理从 Google Earth En-gine(GEE,https://earthengine.google.com)平台收集2022—2023年9—12月份云量少于 5% 的12景Sentinel-2Level-2A 级别影像数据。相关研究表明,森林公园病虫害主要发生于2一12月,高峰期为6一9月,主要表现为染病后叶片衰落,光合、蒸腾作用受到影响,森林的长势削弱,病害发展至9—12月,树上叶片、花果基本落光,枝条枯死[23]。
因上半年云雨天气对影像质量产生较大不利影响。采集影像数据具体时间如下:2022年9月4日、9月14日、10月24日、10月29日、11月13日、12月18日、12月28日,2023年11月18日、11月28日、12月8日、12月23日、12月28日,数据依次经过大气校正、辐射校正、几何校正和云掩膜等处理。
实地调查时间分别为2023年9月8日、10月15日、11月15日、12月7日,实地调查与理论推断一致,主要表现为病害。本研究首先选取病害典型样地,对样地八角林进行调查。采用对角线取样法调查病害树木,每棵树从冠层的东南西北4个方向各选取一组枝条,统计枝条上的叶片数、病斑个数和落叶率。结合“林业有害生物发生与灾害标准LY/T1681—2006”与八角林实际因病害造成的落叶状况,将病害程度划分为4个等级:健康、轻度、中度和重度病害,样本数分别为26、20、23和21。八角病害分级标准见表1,其中,落叶率的计算公式见式(1)[24]。
| 病害等级 | 落叶率 |
| 健康 | 落叶率≤10% |
| 轻度 | 11%≤落叶率≤30% |
| 中度 | 31%≤落叶率≤60% |
| 重度 | 61%≤落叶率 |
1.2.2研究方法流程图如图2所示,主要可分为3个步骤:(1)从GEE平台筛选云量小于 5% 的实验区Sentinel-2图像,并进行预处理;(2)综合分析森林受病虫害胁迫下的形态颜色、结构和功能变化情况,优选植物衰减指数(PSRI)、结构不敏感色素指数(SIPI)、归一化差值红边指数(NDRE)植被光合有效辐射吸收系数(FAPAR)和归一化差值水体指数(NDWI)五个指数作为森林病虫害监测分类特征;(3)选择支持向量机及随机森林算法构建森林病虫害胁迫遥感监测模型,得到2种模型算法的森林病虫害分类结果。1.2.2.1植被病虫害敏感指数选择病虫害会导致植被叶片枯萎、失叶量增高,蒸腾及光合作用能力、水分吸收功能下降、冠层覆盖度减少等形态颜色、功能与结构的变化,进而影响植物的光谱特征[25-26]。根据已有研究,选择对植被颜色变化敏感的植被衰减指数(PSRI),对植被结构变化敏感的结构不敏感色素指数(SIPI)、归一化差值红边指数(NDRE)及对植被功能变化敏感的植被光合有效辐射吸收系数(FAPAR)、归一化差值水体指数(NDWI)5类植被指数开展研究[27-3],这些指数可反映植被色素含量、水分含量及生理代谢活动(呼吸作用和光合作用)的变化情况,可全面评估森林受胁迫状况,具体如表2所示。

1.2.2.2森林病虫害胁迫监测模型通过收集相关的实地数据和遥感影像,构建森林病虫害监测模型,进而识别病虫害发生区域,并评估病害的发生程度。随着机器学习的不断发展,利用机器学习对森林病虫害进行监测,能够处理大量复杂数据,为研究提供更为精细化的监测结果,从而为公园森林资源的可持续管理提供数据支撑。森林病虫害胁迫响应敏感的植被指数见表2。
| 植被指数 | 计算公式 | 说明 |
| 植被衰减指数(PSRI) PSRI= | B4-Bs B2 | 与植被的生长状况和叶绿素含量密切相 关。当植被受到病虫害侵害时,PSRI值 会增加。 |
| 结构不敏感色素指数 SIPI= (SIPI) | Bs-B2 Bg - B4 | 对植被叶片结构变化敏感,可监测植被 健康,检测植物生理胁迫。 |
| 归一化差值红边指数 (NDRE) | Bg - Bs NDRE= Bg + Bs | 对植被叶绿素含量和叶片内部结构等生 理状态敏感性较高,值越大表示植被生 长状况越好。 |
| 植被光合有效辐射吸收系 数(FAPAR) FAPAR= 归一化差值水体指数 (NDWI) | Bs-B4 EVI=2.5x Bg+6×B4-7.5×B+1 | 反映了植被利用光能进行光合作用的能 力,能够有效监测植被生长状况。值变 |
| (EVI-EVI0.02)x(0.95-0.001) + 0.001 | 小则表示植被处于不良环境中或正受病 | |
| EVI0.98-EVI0.02 Bs-B NDWI= Bg + B3 | 虫害攻击。 对植被冠层含水量比较敏感,能够检测 由于受到病虫害导致的水分含量的变 化。 |
(1)支持向量机支持向量机SVM(SupportVectorMachines)是一种按监督学习方式对数据进行二元分类的广义线性分类器,通过选择核函数,在所确定的特征空间上构造分类超平面,采用正则化因子平衡间隔最大化和训练误差最小化的方式实现样本的二分类32]。支持向量机采用核函数的方法解决维数灾难和非线性可分的问题,能够处理病虫害模型中复杂的特征空间,如形态特征、纹理特征和颜色特征等。同时支持向量机适用于小样本数据集。因此,在病虫害样本量较小的情况下,也能够获得良好统计规律。
借助GEE平台强大的遥感数据处理能力,分别计算了2022—2023年每月的5个森林病虫害指数作为模型的分类特征。利用所选择的训练数据集,选择支持向量机作为分类器时,考虑了多种核函数类型,包括线性核函数、多项式核函数和高斯核函数,通过对比实验,发现高斯径向基核函数在森林病虫害分类中的效果较为优越。为了进一步优化模型性能,对不同gamma系数和惩罚项系数c的组合进行了分类精度测试。最终确定,在gamma系数为0.5,惩罚项系数c为1的条件下,分类精度达到最高。这一参数配置不仅提升了分类的准确性,也增强了模型的稳定性和泛化能力。
(2)随机森林算法随机森林(RandomForest,RF)分类算法是一种结合了Bagging和决策树的机器学习算法,是以多棵决策树为基础的集成分类器[33-34]。随机森林能够处理大量的输人变量,且在构建模型时不需要进行特征选择或降维,通过评估每个输入特征的重要性,筛选出有利于识别病虫害的关键特征,实现对病害的准确分类。
利用随机森林分类器,对2022一2023年间六万林场森林公园的森林病虫害信息进行了精细化的分类提取。计算了每个月的5个关键病虫害指数,将其作为模型的分类特征,并对选择的RF分类器进行了详尽的参数优化。通过对比不同数量分类树(即随机森林中的决策树数量)、划分不同比例的训练样本和测试样本的平均精度,发现当随机森林树的数量设定为220,筛选 80% 的样本作为训练样本时,分类误差达到最低。这一参数设定不仅保证了模型的稳健性,同时也优化了模型的性能,使得森林病害信息的提取效果达到最佳。
1.2.2.3精度验证在GEE上随机选择 80% 的样本点用于训练, 20% 的样本点用于验证,采用混淆矩阵,以总体精度和Kappa系数作为评估分类模型精度的评价指标[35]。
总体精度表示所预测分类结果与实际类型相一致的概率,反映了分类结果的正确匹配程度。计算公式见式(2)。
式中, T_{c^{\prime}} T_{N} 表示正确分类的样本数和总样本数。
Kappa系数是一个检测模型预测结果和实际分类结果是否一致的指标,用于衡量分类的效果,Kappa系数的值越大,表示模型的分类准确度越高。计算公式见式(3)。
式中, P_{o} 表示每一类正确分类的样本数量之和除以总样本数,即总体分类精度; P_{e} 是预期的精确一致性的比例,即分类器在随机情况下预测结果与实际结果一致的比例。
2 结果与分析
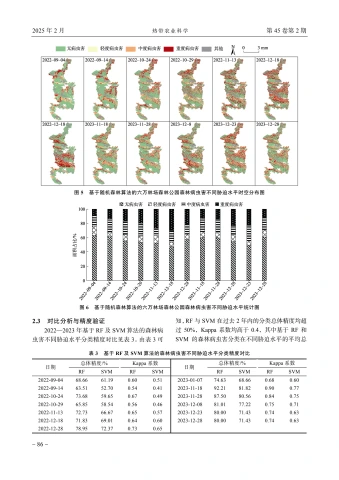
2.1基于支持向量机的森林病虫害遥感信息提取
基于支持向量机的六万林场森林公园森林病虫害不同胁迫水平时空分布及统计见图3、4。六万林场森林公园森林健康状况整体表现良好,大多数区域未受病虫害影响,且在2022一2023年病虫害呈现改善趋势。其中2022年11月13日、12月18日、12月28日,2023年11月28日、12月8日森林病虫害情况相对较为严重;2022年10月24日、10月29日,2023年12月23日、12月28日病虫害情况相对较轻,且病虫害区域呈现集聚分布、连片蔓延的特征,尤其每年11一12月,病虫害蔓延加重趋势更为显著,而后病虫害情况有所缓解。说明六万林场森林公园病虫害易发于每年11一12月,随后治理措施的实施使病虫害得以一定程度控制。从空间来看,六万林场森林公园病虫害程度多介于中度与重度之间,且主要集中于中部山地、丘陵地区,南部地区发生率相对较低,说明研究区森林病虫害严重程度较高,山地、丘陵等地形复杂、人类难以涉足区域,更利于病虫害的发生且难以及时治理和控制。


2.2基于随机森林算法的森林病虫害遥感信息提取
基于随机森林算法的六万林场森林公园森林病虫害不同胁迫水平时空分布如图5所示。由图5可知,2年内研究区森林病虫害多发生于北部与中部地区,较少发生于南部地区,且病虫害胁迫存在明显的规律变化特征,即病虫害分布多聚集、连片,中度、重度占病虫害绝大部分,12月尤为明显,说明研究区受病虫害影响范围较广、程度较深。基于随机森林算法的六万林场森林公园森林病虫害不同胁迫水平统计如图6所示。总体来看,研究区森林生长状况较为良好,均约半数以上未受病虫害侵扰,其中2022年12月18日,2023年12月8日、12月23日森林病虫害比例相对偏高,而2022年9月4日、9月14日,2023年12月28日森林病虫害比例相对偏低,说明12月为研究区病虫害高发期,这与六万林场森林公园森林病虫害高发期相符。而2年内研究区森林病虫害整体呈现变好趋势,说明对病虫害进行及时干预与管控,较为有效地抑制病虫害受灾状况。

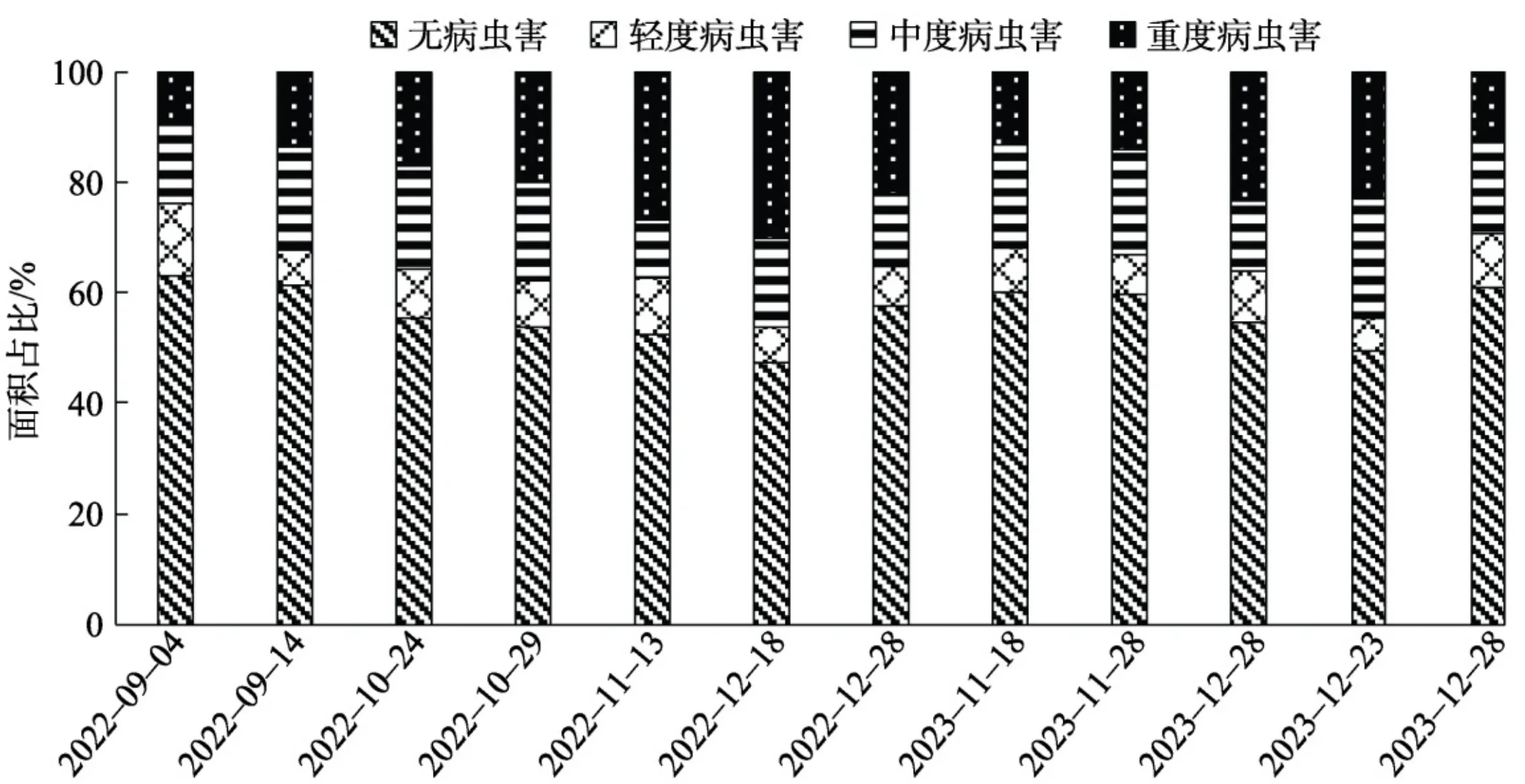
2.3对比分析与精度验证
2022一2023年基于RF及SVM算法的森林病虫害不同胁迫水平分类精度对比见表3。由表3可知,RF与SVM在过去2年内的分类总体精度均超过 50% ,Kappa系数均高于0.4,其中基于RF和SVM的森林病虫害分类在不同胁迫水平的平均总体精度分别为 76.20% 68.56% ,平均Kappa系数分别为0.70、0.60,且基于RF的分类效果在每一时期均优于SVM,说明RF及SVM算法均具备一定的病虫害监测能力,但RF方法的优势更为显著。
| 日期 | 总体精度/% | Kappa系数 | 日期 | 总体精度/% | Kappa系数 | ||||
| RF | SVM | RF | SVM | RF | SVM | RF | SVM | ||
| 2022-09-04 | 68.66 | 61.19 | 0.60 | 0.51 | 2023-01-07 | 74.63 | 68.66 | 0.68 | 0.60 |
| 2022-09-14 | 63.51 | 52.70 | 0.54 | 0.41 | 2023-11-18 | 92.21 | 81.82 | 0.90 | 0.77 |
| 2022-10-24 | 73.68 | 59.65 | 0.67 | 0.49 | 2023-11-28 | 87.50 | 80.56 | 0.84 | 0.75 |
| 2022-10-29 | 65.85 | 58.54 | 0.56 | 0.46 | 2023-12-08 | 81.01 | 77.22 | 0.75 | 0.71 |
| 2022-11-13 | 72.73 | 66.67 | 0.65 | 0.57 | 2023-12-23 | 80.00 | 71.43 | 0.74 | 0.63 |
| 2022-12-18 | 71.83 | 69.01 | 0.64 | 0.60 | 2023-12-28 | 80.00 | 71.43 | 0.74 | 0.63 |
| 2022-12-28 | 78.95 | 72.37 | 0.73 | 0.65 | |||||
3 讨论与结论
森林病虫害的发生不仅会改变植被的形态、颜色等外部特征,还会对光合作用、蒸腾作用等生理功能产生影响[36]。本研究选取综合表征植被颜色、结构、功能变化的植被指数,基于支持向量机和随机森林等机器学习算法构建森林病虫害遥感监测模型。结果表明,森林病虫害具有聚集和连片蔓延的特点,且随机森林算法构建的森林病虫害遥感监测模型要优于支持向量机算法,这与前人的研究结果一致[37-38]。
尽管构建的森林病虫害遥感监测模型取得了较好的效果,但研究仍存在一定的局限性。首先,病虫害、营养、干旱等多种胁迫往往混杂在一起[39],综合植被颜色、结构和功能变化的植被指数可一定程度反映病虫害胁迫特征,但未考虑与病虫害胁迫特征相似的其他胁迫因素影响。在今后研究中,将进一步探索病虫害胁迫与其他不同胁迫的光谱响应特征,以提高森林病虫害胁迫识别的准确性。此外,研究仅对比随机森林与支持向量机两类机器学习算法在森林病虫害监测中的效果,但其他的机器学习方法在森林病虫害遥感监测中的效果有待进一步验证。
为了有效地监测植被病虫害动态变化,迫切需要高时空分辨率的遥感数据。但云雨天气的影响限制了光学遥感影像的使用。未来发展SAR与光学遥感融合算法,生成同时具有高时空分辨率的遥感影像,是森林病虫害遥感分析的基础。
本研究探讨病虫害胁迫的森林光谱特征,优选可表征森林病虫害胁迫程度的光谱指标,以RF及SVM算法分别建立森林病虫害胁迫遥感监测模型,主要结论如下。
(1)综合表征森林植被颜色、结构、功能变化的PSRI、SIPI、NDRE、FAPAR、NDWI 指数可有效监测森林病虫害不同胁迫水平。
(2)RF及SVM算法均具备一定的森林病虫 害监测能力,总体分类精度分别为 63.51%~ 92.21% 、 52.70%~81.82% ,Kappa系数分别为
0.54\~0.90、0.41\~0.77。总体来看,RF分类效果好于SVM分类效果,更有利于森林病虫害的监测。
(3)广西六万林场森林公园2022和2023年的12月病虫害最为严重,中度、重度区占比最大,主要集中于森林公园中部高山与北部丘陵地带,且多为山的北坡,同时呈现明显聚集、连片蔓延的分布特征。
参考文献
[1]王兵,任晓旭,胡文.中国森林生态系统服务功能及其价 值评估[J].林业科学,2011,47(2):145-153.
[2]孙伟韬.基于多光谱遥感影像的森林病虫害监测研究[]. 按树科技,2023,40(4):39-46.
[3]吴霜,延晓冬,张丽娟.中国森林生态系统能值与服务功 能价值的关系[叮].地理学报,2014,69(3):334-342.
[4] Choi W I, Park Y S. Monitoring, assessment and management of forest insect pests and diseases[J]. Forests, 2019, 10(10): 865.
[5]Heuss M, D'Amato A W, Dodds K J. Northward expansion of southern pine beetle generates significant alterations to forest structure and composition of globally rare Pinus rigida forests[J]. Forest Ecology and Management, 2019, 434: 119-130.
[6]Fernandez-Carrillo A, Patocka Z, Dobrovolny L, et al. Monitoring bark beetle forest damage in central Europe.A remote sensing approach validated with field data[J]. Remote Sensing, 2020, 12(21): 3 634.
[7]Abdullah H, Darvishzadeh R, Skidmore A K, et al. Sensitivity of landsat-8 OLI and TIRS data to foliar properties of early stage bark beetle (Ips typographus L.) infestation[J]. Remote Sensing, 2019, 11(4): 398.
[8]黄旭影,许章华,林璐,等.基于毛竹叶片理化参数的刚 竹毒蛾危害检测研究[J].光谱学与光谱分析,2019,39(3): 857-864.
[9]竞霞,邹琴,白宗,等.基于反射光谱和叶绿素荧光数 据的作物病害遥感监测研究进展[]].作物学报,2021, 47(11): 2 067-2 079.
[10] Abdullah H, Skidmore A K, Darvishzadeh R, et al. Sentinel-2 accurately maps green-attack stage of European spruce bark beetle (Ips typographus L.) compared with Landsat-8[J]. Remote Sensing in Ecology and Conservation, 2019, 5(1): 87-106.
[11] Xu Z H, Huang X Y, Lin L, et al. BP neural networks and random forest models to detect damage by Dendrolimus punctatus Walker[J]. Journal of Forestry Research, 2020, 31(1): 107-121.
[12]郭仲伟,吴朝阳,汪箫悦.基于卫星遥感数据的森林病虫 害监测与评价[叮].地理研究,2019,38(4):831-843.
[13] Zhang Y H, Dian YY, Zhou J J, et al. Characterizing spatial patterns of pine wood nematode outbreaks in subtropical zone in China[J]. Remote Sensing, 2021, 13(22): 4 682.
[14]Lei S H, Luo JB, Tao X J, et al. Remote sensing detecting of yellow leaf disease of arecanut based on UAV multisource sensors[J]. Remote Sensing, 2021, 13(22): 4 562.
[15]彭隆赞,金时超,孟凡凡,等.马尾松毛虫危害程度的高 光谱监测方法[].湖北林业科技,2017,46(3):1-6.
[16]赵晋陵,金玉,叶回春,等.基于无人机多光谱影像的槟 榔黄化病遥感监测[J].农业工程学报,2020,36(8):54-61.
[17]Luo Y Q, Huang HG, Roques A. Early monitoring of forest wood-boring pests with remote sensing[J]. Annual Review of Entomology, 2023,68: 277-298.
[18]许章华,黄旭影,林璐,等.基于Fisher 判别分析与随机 森林的马尾松毛虫害检测[J.光谱学与光谱分析,2018, 38(9): 2 888.
[19]黄芳芳,雷鸣,张力,等.基于随机森林和决策树的马尾 松松材线虫病监测方法[]].信息通信,2019,32(12):32-36.
[20] Zhan Z Y, Yu L F, Li Z, et al. Combining GF-2 and sentinel-2 images to detect tree mortality caused by red turpentine beetle during the early outbreak stage in North China[J]. Forests, 2020, 11(2): 172.
[21] Xu Z H, Zhang Q, Xiang S Y, et al. Monitoring the severity of Pantana phyllostachysae Chao infestation in mosobamboo forests based on UAV multi-spectral remote sensing feature selection[J]. Forests, 2022, 13(3): 418.
[22]黄云.广西按树病虫害的发生现状与防治措施[].南方农 业, 2023, 17(12): 76-78.
[23]周云飞.八角炭疽病的发生特征与防治措施[].乡村科 技,2021,12(5):80-81.
[24]蒋晓萍,莫小刚,周婵,等.广西八角炭疽病年发病节律 观测与防治适期分析[].中国野生植物资源,2017,36(6): 68-70+79.
[25]李学平.小型无人机林间施药防治八角炭疽病效果试验 [].林业科技通讯,2019(9):49-50.
[26]莫小刚.八角炭疽病发病原因及防治措施探析[].南方农 业, 2021, 15(2): 62-63.
[27] Merzlyak M N, Gitelson A A, Chivkunova O B, et al. Non-destructive optical detection of pigment changes during leaf senescence and fruit ripening[J]. Physiologia Plantarum, 1999, 106(1): 135-141.
[28] Haboudane D, Miller J R, Tremblay N, et al. Integrated narrow-band vegetation indices for prediction of crop chlorophyll content for application to precision agriculture[J]. Remote Sensing of Environment, 2002, 81(2/3): 416-426.
[29]Shu M Y, Gu X H, Zhou L F, et al. Establishing NDRE dynamic models of winter wheat under multi-nitrogen rates based on a field spectral sensor[J]. Applied Optics, 2021, 60(4): 993-1 002.
[30] Li L, Du Y M, Tang Y, et al. A new algorithm of the FPAR product in the Heihe river basin considering the contributions of direct and diffuse solar radiation separately[J]. Remote Sensing, 2015, 7(5): 6 414-6 432.
[31]McFeeters S K. The use of the Normalized Difference Water Index (NDWI) in the delineation of open water features[J]. International Journal of Remote Sensing, 1996, 17(7): 1 425-1 432.
[32]杨亚玲.基于TM遥感影像的河套灌区土地利用变化分析 与实现[D].杨凌:西北农林科技大学,2017.
[33]闫东阳.基于对象的随机森林遥感分类方法优化[D].北 京:中国地质大学(北京),2018.
[34]王猛,张新长,王家耀,等.结合随机森林面向对象的森 林资源分类[J].测绘学报,2020,49(2):235-244.
[35]肖庆琳,张加龙,曹军,等.耦合多特征多时相的普洱市 优势树种分类研究[].森林工程,2024,40(2):117-126.
[36]Abd El-Ghany N M, Abd El-Aziz S E, Marei S S. A review: Application of remote sensing as a promising strategy for insect pests and diseases management[]. Environmental Science and Pollution Research International, 2020, 27(27): 33 503-33 515.
[37] Migas-Mazur R, Kycko M, Zwijacz-Kozica T,et al. Assessment of sentinel-2 images, support vector machines and change detection algorithms for bark beetle outbreaks mapping in the Tatra mountains[J]l. Remote Sensing, 2021, 13(16): 3 314.
[38] Sothe C, De Almeida C M, Schimalski M B, et al. Comparative performance of convolutional neural network, weighted and conventional support vector machine and random forest for classifying tree species using hyperspectral and photogrammetric data[J]. GIScience & Remote Sensing, 2020, 57(3): 369-394.
[39] Wang S Y, Liu M L, Li Y M, et al. Spatiotemporal cube model based on stress features for identification of heavy metal stress in rice[J]. IEEE Transactions on Geoscience and Remote Sensing, 2024, 62: 4 401 313.
金钱树种质资源倍性鉴定
周成诚1.2 陆锦萍」陈金花」牛俊海」张香秦’冷青云{(1.中国热带农业科学院热带作物品种资源研究所/农业农村部华南作物基因资源与种质创制重点开放实验室/海南省热带观赏植物种质创新利用工程技术研究中心/海南省热带作物资源遗传改良与创新重点实验室海南海口571101;2.福建农林大学福建福州350002;3.乐东普英洲花卉园艺有限公司海南乐东572541)
摘要金钱树是一种优良的室内观叶植物,随着该产业的发展,培育新品种和种质资源评价尤为重要。倍性鉴定是花卉种质资源鉴定及新品种选育的重要工作之一。研究以4个金钱树品种为试验材料,采用形态学特征比较及流式细胞术测定对其进行鉴定。结果表明:粗杆品种叶片厚度最厚,叶序轴粗度最大,与其他品种差异显著。粗杆的气孔密度与其他3个品种存在显著差异,其气孔密度最小,为18.52个 /mm^{2} ;粗杆的叶片单细胞DNA含量是其他3个品种的2倍,为四倍体植株。本研究明确了圆叶、尖叶及黑钻3个品种为二倍体植株,粗杆植株为四倍体植株,为金钱树新种质培育提供重要依据。
关键词金钱树;种质资源;倍性鉴定
32.36 文献标识码A DOI: 10.12008/j.issn.1009-2196.20
Ploidy Identification of Zamioculcas zamiifolia
ZHOU Chengchengl2 LU Jinping' CHEN Jinhua' NIU Junhai' ZHANG Xiangqin? LENG Qingyun'
(1.Tropical Crop Germplasm Research Institute, Chinese Academy of Tropical Agricultural Sciences/Key Open Laboratory of Crop Gene Resources and Germplasm Creation in South China, Ministry of Agriculture and Rural Affairs/Engineering Technology Research Center for Innovative Utilization of Tropical Ornamental Plant Germplasm in Hainan Province/Key Laboratory of Genetic Improvement and Innovation of Tropical Crop Resources in Hainan Province,Haikou,Hainan 571101, China; 2.Fujian Agriculture and Forestry University,Fuzhou,Fujian 350002, China; 3. Ledong Puyingzhou Flower and Horticulture Co.,Ltd.,Ledong, Hainan 572541, China)
Abstract Zamioculcas zamifolia is an excellent indoor foliage plant, and with the development of this industry, the cultivation of new cultivars and the evaluation of germplasm resources are crucial. Ploidy identification is one of the vital tasks in the identification of germplasm resources and the breeding of new varieties of flowers.Four cultivars of Z. zamifolia were used as test materials, their morphological characteristics were compared, and flow cytometry was used for detection.The results revealed that the^Coarse Rod’cultivar had the thickest leaves and the thickest petioles,which significantly differedfrom those of the other cultivars. The stomatal density of the^Coarse Rod’ cultivar was significantly different from that of the other three cultivars,and the stomatal density was the lowest(18.52individuals *mm^{-2} ),and the single-cellDNA content of theleavesof ^Coarse Rod’was twice that of the other three cultivars, indicating that it is a tetraploid plant. In this study, it was confirmed that the three cultivars, “Round Leaf",“Sharp Leaf", and “Black Diamond',were confirmed to be diploid plants, and the Coarse Rod’ plants were tetraploids, providing a vital basis for the cultivation of new germplasms.
Keywords Zamioculcas zamifolia; germplasm resources; ploidy identification
金钱树(Zamioculcaszamifolia)为天南星科(Aroceae)雪铁芋(ZamioculcasHW.Schott)属多年生常绿草本植物,原产地为热带非洲东部坦桑尼亚,其株型美观,茎的基部膨大成球形,羽状复叶,小叶肉质、椭圆,叶色墨绿,富有光泽,整个复叶像一串串铜钱",为优良的观叶植物,也可作布景、点缀园林配置应用,是花卉市场上的热销品种之一。金钱树自1997年引人中国,有近30年的栽培历史,在研究方面主要集中在组织培养、种苗繁育技术及栽培技术研究等领域2-5],在种质资源评价及新品种培育等方面报道较少。目前,市场上流通的金钱树品种为普通尖叶品种,极为单一,随着金钱树产业的发展,对新品种的需求日益迫切,培育适宜我国热区气候和生产条件的品种是该产业发展的必然之路。对收集的金钱树种质进行评价尤为关键,其中,倍性鉴定是种质资源评价的重要内容。植物倍性鉴定在植物遗传育种中具有重要作用,利用倍性鉴定,可以快速筛选出多倍体植株,大大减少育种的盲目性及工作量,加速育种进程。
倍性鉴定方法有形态特征观察、染色体计数鉴定、分子水平鉴定、流式细胞术鉴定等[7-]。其中,形态特征观察主要从器官形态学特征、气孔大小等方面鉴定倍性,该方法受材料生理状态影响较大,准确性不高。染色体计数法虽准确度高,但对细胞学操作技术要求较高,且费时费工,流式细胞术在植物研究中起步较晚,但由于它具有制样简单、灵敏度及准确性较高、数据的可重复性等优势,已逐渐发展为一种常用的植物倍性和基因组大小预估的研究方法[11-16]。本研究以 4个金钱树品种为材料,采用形态学特征比较、流式细胞术检测进行倍性鉴定,旨在明确不同品种金钱树倍性水平,为其新种质培育提供依据。
1 材料与方法
1.1材料
供试材料为中国热带农业科学院热带作物品种资源研究所海南省热带草本花卉种质资源圃(儋州)种植的金钱树品种尖叶、黑钻、圆叶及粗杆植株,引种自乐东普英洲花卉园艺有限公司。
1.2方法
1.2.1外观形态特征比较取4个品种的植株, 测量植株高度、叶片长度、叶片宽度、叶间距、 叶片厚度、叶序轴粗度。
1.2.2叶片气孔大小和密度比较参照秦卫华的方法[],取4个金钱树品种成熟叶片,洗净后选取叶片中间部分去除叶脉后用锋利的刀片裁剪成大小一致的小块,面积约为 2\;cm{x}2\;cm 5% NaOH浸泡处理 7~8~h~ ,用清水漂洗干净后,用镊子撕取叶脉外的叶下表皮制片观察。用镊子撕下其下表皮置于载玻片上,载玻片上滴有去离子水,用滤纸吸取多余的水分,加盖玻片后放置于生物显微镜下观察测量(显微镜型号为Phenix凤凰PH100)。每个品种至少观察5个视野,记录每个视野内气孔器的个数,每张制片选取20个气孔,测量每个气孔器的长度和宽度。
1.2.3流式细胞仪检测参照 Joao L及冷青云[8-19]的方法,取4个品种植株幼嫩叶片 0.01 ~g~ ,置于含 1~mL WPB缓冲液的培养皿中用刀片切碎,WPB缓冲液的配方为( 0.2 \mol/L Tris-HCl, 2\;\mmol/L EDTA -Na_{2*2H_{2O},4\mm o l/L\ M a C l_{2}-6H_{2O},86\mm o l/L} {NaCl} , 1% PVP-10, 1% (V/V)Triton X-100,10\mmol/L 焦亚硫酸钠, pH~7.5 ), 1 \min 后用尼龙网(300目)过滤,滤液加人 100\ \upmuL\ 10\ mg/mL DAPI, 2~min 后待染液与过滤液充分混匀上机检测,仪器型号为 SysmexCyFlow \ensuremath{\mathbb{R}} Ploidy Analyser,激发光为 488~nm ,每次检测收集5000个颗粒,变异系数控制在 5% 以内。每份材料重复3次。
2 结果与分析
2.1金钱树4个品种的形态学特征比较
金钱树粗杆品种较其他3个品种(图1-A\~1-C)植株矮小,株型更紧凑(图1-D),叶形更大(图1-E),叶序轴更短、更粗(图1-F)。4个金钱树品种的形态特征比较见表1。在植株高度上,金钱树尖叶品种与其他3个品种均存在显著差异,且株高最大,为 80.32~cm ;粗杆品种株高最小,但与圆叶品种差异不显著。在叶片长度上,粗杆品种叶长最大,与黑钻和圆叶品种存在显著差异,但与尖叶品种差异不显著。圆叶品种的叶片宽度最大,为 6.54~cm ,与其他3个品种存在显著差异。圆叶品种的叶长与叶宽之比最小,为1.52,与其他3个品种均存在显著差异。黑钻品种的叶间距低于其他3个品种,且存在显著差异。粗杆品种的叶片厚度和叶序轴粗度均高于其他3个品种,且存在显著差异。

A.尖叶;B.黑钻;C.圆叶;D.粗杆;E.叶序轴形态(a.尖叶;b.黑钻;c.圆叶;d.粗杆);F.叶片形态(a.尖叶;b.黑钻;c.圆叶;d.粗杆)
2.2金钱树4个品种的气孔特征比较
金钱树4个品种的气孔特征比较见图2。从表2可看出,金钱树粗杆品种的气孔长度为43.31~{\upmum} ,大于其他3个品种,且存在显著差异。粗杆品种的气孔密度也与其他3个品种存在显著差异,气孔密度最小,为18.52个 /mm^{2} 。圆叶品种的气孔宽度最大,为 10.50~{\upmum} 。4个品种的气孔宽度差异不显著。
2.3金钱树4个品种的DNA含量分析
由叶片单细胞DNA含量分布(图3)可看出,尖叶、黑钻及圆叶3个品种的峰值在荧光强度为6250通道的位置,而粗杆品种的峰值在荧光强度为12500通道的位置,由此可看出,粗杆品种叶片单细胞DNA含量是其他3个品种的2倍。这表明圆叶、尖叶及黑钻3个品种为二倍体,粗杆品种为四倍体。
3 讨论与结论
金钱树因其较高的观赏价值,被广泛商业种植。然目前市场主栽的品种仅为尖叶1种,品种极为单一,这与金钱树的物种生物特性有关系。金钱树原产地在非洲的坦桑尼亚等地,为单属单种植物,引种到国内后,环境与原产地差异较大,结实率非常低,未见到过其自然结实的种子,另外金钱树在原产地也需人工授粉才能得到种子,种子发芽率也较低,从种子到成株需要3\~5年时间,这在很大程度上限制了其育种,相关育种技术积累有限。金钱树的繁殖主要是以无性繁殖为主,其中以叶插法繁殖最为普遍,也是目前商业种植使用的最主要的手段,在大量且多代的叶插繁殖过程中也会有一定的变异,本研究中评价的圆叶和粗杆2个品种就是在商业种植基地发现并培育的,在很多花卉中均有利用芽变培育成新品种,如月季、菊花、兰花、红掌等[19-2]。利用秋水仙素处理无菌材料或者块茎也可以获得一些多倍体植物变异[23-25]。因此,发现变异、诱导产生变异、利用变异不能正常有性结实,而是只能以无性繁殖为主,这也是花卉培育新品种行之有效的手段。
| 品种 | 株高/cm | 叶长/cm | 叶宽/cm | 叶间距/cm | 叶片厚度/mm | 叶序轴粗/mm | 叶长/叶宽 |
| 尖叶 | (80.32±8.18) a | (11.12±0.77) ab | (5.30±0.68)b | (6.40±0.67) a | (0.70±0.06)c | (26.61±7.08) bc | (2.11±0.13) a |
| 黑钻 | (44.89±4.11) c | (8.99±0.99) c | (4.04±0.47) c | (3.24±0.99) c | (1.02±0.00) b | (21.62±1.86) c | (2.23±0.22) a |
| 圆叶 | (57.88±1.83) b | (9.92±0.91) bc | (6.54±0.27) a | (4.98±0.55) b | (0.76±0.08)c | (27.57±2.53) bc | (1.52±0.10) b |
| 粗杆 | (51.66±2.04) b | (12.22±1.46) a | (5.42±0.17) b | (5.70±1.29) ab | (1.32±0.06) a | (29.46±1.37) a | (2.26±0.31) a |
图2金钱树4个品种的气孔特征比较
| 品种 | 气孔长度/um | 气孔宽度/um | 气孔密度/(个·mm-2) |
| 尖叶 | (28.74±5.42) b | (8.26±2.38) a | (36.49±3.82) a |
| 黑钻 | (33.04±2.62) b | (8.19±2.90) a | (25.05±2.67) b |
| 圆叶 | (27.65±2.27) b | (10.50±3.57) a | (25.05±1.69)b |
| 粗杆 | (43.31±6.61) a | (9.47±3.78) a | (18.52±2.67)c |

A.尖叶;B.黑钻;C.圆叶;D.粗杆。
多倍体育种是观赏植物育种的重要方向之一。与二倍体相比,多倍体植株在外部形态上会有所改变,如叶片厚度增加、叶色加深、气孔变大、气孔密度降低、花粉粒变大、育性和抗性改变等[26-28]。外部形态变化可以作为初步筛选多倍体的方法,紫薇、红掌、兰花等多倍体植株的鉴定,形态学特征就作为一个鉴定指标9,19,29。本试验结果中粗杆品种叶片厚度最厚,气孔长度增大,密度降低,与其他品种差异显著,也表明外部形态特征也可以作为金钱树多倍体初筛的重要参数。
单靠形态变异来确定植物倍性是不够准确的,需要进一步倍性鉴定,流式细胞术检测是植株倍性水平可靠的方法。在紫薇、鹤顶兰等多倍体鉴定中发现,变异株有一定比例为非多倍体或者混倍体[29-30]。本试验材料经流式细胞仪检测,粗杆品种DNA含量为其他品种的2倍,确定为四倍体,未发现混倍体的存在。本研究通过植株外部形态进行初筛,进一步通过流式细胞术确定倍性,这对金钱树多倍体的鉴定是可行的,也为金钱树多倍体育种倍性鉴定提供可靠方法。
参考文献
[1]徐世松,黄少华,黄素荣,等.海南金钱树设施栽培技术[].热带农业工程,2009,33(1):69-71+79.
[2]王建宇,孙兆军.北方金钱树繁育技术研究[].北方园艺,2007(4): 127-128.
[3]谢春艳,张国武,胡松竹,等.不同轻型基质对金钱树生长影响初报[J.热带作物学报,2010,31(3):365-370.
[4]林茂,王华新,唐遒冥,等.金钱树组织培养研究进展[]].广东农业科学,2012,39(24):56-58+68.
[5]康冬茹,苑晓亮,赵晓丹,等.金钱树叶片再生体系的建立[J].山东林业科技,2013,43(2):63-65.
[6]杨清淮.植物染色体倍性鉴定方法概述[].现代园艺,2014(3): 12-13.
[7]陶抵辉,李小红,王利群,等.植物染色体倍性鉴定方法研究进展[J.生命科学研究,2009,13(5):453-458.
[8]张虹,路国栋,赵帅,等.利用流式细胞仪鉴定植物倍性的研讨[J].分子植物育种,2020,18(11):3713-3717.
[9]冷青云,陆锦萍,黄少华,等.基于流式细胞术的兰属杂交后代倍性鉴定[J].热带作物学报,2023,44(11):2219-2 226.
[10]田新民,周香艳,弓娜.流式细胞术在植物学研究中的应用—检测植物核DNA含量和倍性水平[J.中国农学通报,2011, 27(9): 21-27.
[11]张志珂,王永清,林顺权,等.借助细胞流式仪进行枇杷基因组学测序材料的倍性鉴定[J].果树学报,2012,29(3):498-504.
[12]Favoreto C F.,Carvalho R C.,Lima P B A.,et al.Genomesize and base composition of Bromeliaceae species assessedby flow cytometry[J]. Plant Systematics and Evolution, 2012,298(6): 1 185-1 193.
[13] Sayantani N, Kumar S M, Sumita J. An improved method ofgenome size estimation by flow cytometry in five mucilagi-nous species of Hyacinthaceae. [J]. Cytometry. Part A: thejournal of the International Society for Analytical Cytology,2014, 85(10): 833-840.
[14]张宁宁,杨静,孙卫邦.利用流式细胞仪测定荚属植物的基因组大小(英文).植物分类与资源学报,2014,36(6): 730-736.
[15]柳觐,李开雄,孔广红,等.云南芒果种质基因组大小测定与变异分析[J].热带亚热带植物学报,2015,23(4):386-390.
[16]杨转英,吴传龙,丰锋,等.不同株系菠萝蜜染色体倍性及基因组大小分析[J].果树学报,2015,32(4):567-571.
[17]秦卫华,汪恒英,周守标.植物叶表皮永久制片技术的改进[J].生物学杂志,2003(3):38-41.
[18] Joao L, Eleazar R, Jaroslav D, et al. Two new nuclear isola-tion buffers for plant DNA flow cytometry: a test with 37species.[J]. Annals of botany, 2007, 100(4): 875-888.
[19]冷青云,牛俊海,尹俊梅,等.红掌品种^樱桃红'变异株的倍性鉴定[J].热带作物学报,2014,35(11):2139-2143.
[20]周厚高,王文通,乔志钦,等.切花小菊新品种"缤纷"[].园艺学报,2015,42(1):201-202.
[21]金建鹏,高洁,谢琦,等.墨兰新品种^四季花"[J].园艺学报,2022,49(S1):127-128.
[22]汪有良,孙海楠.《ModernRose 12》登录的月季品种群体中自然芽变现象分析[.江苏林业科技,2022,49(4):31-36.
[23]梁君.金钱树(Zamioculcas zamifoliaRehd.)快繁和四倍体新种质选育研究[D].重庆:西南大学,2008.
[24] Vanzie-Canton, D S, Leonhardt., et al. In vitro callus induc-tion and plantlet regeneration protocol dev-eloped for theoryzalin treatment of Zamioculcas zamifolia (Lodd.) Engl.(Araceae).[J]. Acta Horticulturae, 2009, 2009(813): 201-207.
[25]谢添伟,莫饶,唐燕琼,等.金钱树四倍体的诱导与鉴定[J].热带农业科学,2012,32(4):5-9.
[26]Liu G, Li Z, Bao M. Colchicine-induced chromosome dou-bling in Platanus acerifolia and its effect on plant morphol-ogy[J]. Euphytica, 2007, 157(1-2): 145-154.
[27] Tang Z, Chen D, Song Z, et al. In vitro induction and identi-fication of tetraploid plants of Paulownia tomentosa[J]. PlantCell, Tissue and Organ Culture (PCTOC), 2010, 102(2):213-220.
[28]李淑洁,裴怀弟,刘新星,等.植物染色体加倍的表型和遗传学效应研究进展[J].寒旱农业科学,2023,2(12):1 079-1 084.
[29] Zhang X, Hu C,Yao J. Tetraploidization of diploid Dio-scorea results in activation of the antioxidant defense systemand increased heat tolerance[J]. Journal of Plant Physiology,2009, 167(2): 88-94.
[30]李豆豆,刘芝龙,黄明忠,等.鹤顶兰四倍体植株的诱导与鉴定[J].园艺学报,2013,40(10):2033-2038.
基于主成分分析法的15个秋石斛品种综合评价
梁昕景 李香 符洁 邢艳冰 张孟锦(三亚市林业科学研究院海南三亚572000)
摘要为了解誓言等15个秋石斛品种的形态学性状,利用主成分分析法对誓言等15个秋石斛品种进行观察和测量,并进行综合评价。结果表明,品种誓言的花朵最大,泼墨的最小;红霞的花序梗最长,水晶的最短;红金珠花序梗最粗,水晶的最细;贵妃的花梗最长,小男孩的最短;贵妃的花梗最粗,彩霞的最细;彩霞的单株花朵数最多;红金珠和泼墨的花色得分最高,金如意的最低。所测的大多数性状显著相关。22个形态学性状可以转化为6个主成分,主成分累计贡献率为 83.97% ,主成分综合得分排名第1的为誓言,其次为泼墨、小男孩。
关键词秋石斛;品种;主成分分析;综合评价 中图分类号S682.31 文献标识码 A DOI: 10.12008/j.issn.1009-2196.2025.02.015
Comprehensive Evaluation of 15 Dendrobium hybridus Cultivars via Principal Component Analysis
LIANG Xinjing LI Xiang FU Jie XING Yanbing ZHANG Mengjin (Sanya Academy ofForestry,Sanya,Hainan 572000,China)
Abstract To understand the morphological characteristics, the morphological characteristics of 15 Dendrobium hybridus cultivars, e.g., Shiyan, were observed and measured, and principal component analysis was used for comprehensive evaluation. The results revealed that the variety ^Shiyan’had the largest flowers and “Enobi Purple Splash’had the smallest flowers. The inflorescence stem of“Hongxia’is the longest, whereas the^Crystal’ stem is the shortest. The inflorescence stem of the “Hongjinzhu'is the thickest, and ^Crystal’ is the thinnest. The flower stalk of“Guifei'is the longest, while that of“Xiaonanhai' is the shortest. The flower stalk of“Guifei’is the thickest, while^Caixia’is the thinnest.^Caixia’has the highest number of flowers per plant. The highest score is for the colors of “Hongjinzhu’and “Enobi Purple Splash',whereas^Jinruyi’' had the lowest score. Most of the measured traits were significantly correlated. Twenty-two morphological characteristics can be transformed into six principal components, with a cumulative contribution rate of 83.97% .The variety with the highest overall principal component score is Shiyan, followed by‘Enobi Purple Splash’and Xiaonanhai.
Keywords Dendrobium hybridus; cultivars; principal component analysis; comprehensive evaluation
石斛属(Dendrodium.)是多年生草本植物,兰科中三大属之一,全球原生种有约1500种,绝大多数为附生生长,栽培品种一般分为春石斛和秋石斛两大类。在我国,石斛兰是深受消费者喜爱的花卉,近年来育种工作受到重视,已培育出多个新品种[-4。秋石斛较春石斛更耐高温,适合在海南热带地区种植。开展品种资源的性状评价对优良品种筛选与开发以及育种具有重要的意义,秋石斛已有较多相关研究-7。
品种资源评价的方法较多,如,武荣花?利用模糊数学法、李静利用Q型聚类分析法对石斛属资源进行了研究与评价。主成分分析法是应用比较广泛的综合评价方法之一,主要是将多个指标转化为少数几个综合指标进行评价,该方法已在玉米[10]、马铃薯[、甜瓜[2]、和菊芋[13]等多种作物上得到了应用。本研究以搜集的15种生长周期一致的秋石解品种为供试材料,在相同栽培条件下测定品种多个性状指标,并采用主成分分析法进行综合评价,为优良品种和育种亲本筛选提供参考和依据。
材料与方法
1.1材料
供试15个石斛兰栽培品种名称、编号、来源情况见表1,品种性状见图1。
1.2方法
1.2.1试验设计试验在海南省三亚市天涯区三亚市林业科学研究院国家兰花种质资源圃内进行。栽培在智能连栋塑料大棚内。每个品种种植16株,使用水苔盆栽种植,花盆直径3寸(1寸 \approx 3.33~cm ),按常规商品种种植方法进行水肥管理。1.2.2指标测定随机选择3株开花的石斛兰植株测量各项指标,测量使用仪器:常规直尺、数显游标卡尺、国际通用的英国皇家园艺学会比色卡( Royal Horticultural Society Colour Chart, 2015年版)。开花期测量中部成熟叶片长宽,叶片长为从叶顶端边缘到叶基部边缘的最大距离;叶片宽度为与叶片主脉垂直方向最宽处,每片叶测量一次;花朵相关性状的测量和观察参照《植物品种特异性、一致性和稳定性测试指南一石斛属》方法。
| 编号 | 品种 | 拉丁名 | 供苗单位 | 编号 | 品种 | 拉丁名 | 供苗单位 |
| A | 朝日 | / | 五指山万泉园艺有限公司 | 一 | 红玫瑰 | / | 五指山万泉园艺有限公司 |
| B | 黄金富贵 | / | 五指山万泉园艺有限公司 | J | 彩霞 | / | 五指山万泉园艺有限公司 |
| C | 贵妃 | / | 五指山方泉园艺有限公司 | K | 宝石 | / | 五指山万泉园艺有限公司 |
| D | 誓言 | 五指山万泉园艺有限公司 | L | 红霞 | 五指山万泉园艺有限公司 | ||
| E | 金如意 | / | 五指山万泉园艺有限公司 | M | 泼墨石斛 | DendrobiumEnobi PurpleSplash | 五指山万泉园艺有限公司 |
| F | 东方小姐 | / | 五指山万泉园艺有限公司 | N | 小男孩 | / | 五指山万泉园艺有限公司 |
| G | 红金珠 | / | 五指山万泉园艺有限公司 | 0 | 胭脂 | 五指山万泉园艺有限公司 | |
| H | 水晶 | / | 五指山万泉园艺有限公司 |

1.2.3数据统计与分析采用Excel2019对试验数据进行整理,利用SPSS26.0软件对15份石斛兰品种资源22个农艺学性状进行主成分分析,在主成分分析中,按照特征值大于1.0的原则确定主成分个数,并根据特征向量值划分各个性状在主成分中的归属[4];差异性检验采用Duncan's新复极差法进行多重比较。
2 结果与分析
2.1不同石斛兰品种植株大小比较
石斛兰植株长势是进行商品化选择的一个重要因素。由表2可知,15个石斛兰品种假鳞茎长度为 21.33~59.67\mm ,最长的品种为黄金富贵,显著高于其他品种,最短者为水晶,显著低于其他品种;茎粗为 11.87~19.10\;mm ,最粗者为誓言,与多数品种差异不显著,最细者为金如意,显著低于其他品种;植株高度为 129.33~392.33~mm ,黄金富贵最高,其次为誓言、胭脂,泼墨石斛最低,其次为小男孩、水晶,显著低于其他品种;植株宽度为 181.73~297.67\mm ,誓言最宽,其次为红霞、贵妃、彩霞,黄金富贵最窄,且显著低于大部分品种;叶片长度为 86.33~163.73\mm ,红霞最长,其次为誓言、贵妃,水晶最短,再次为泼墨石斛,显著低于其他品种;叶片宽度为 28.27~ 66.10~mm ,誓言最宽,显著高于其他品种,水晶最窄,其次为宝石、朝日。
| 序号 | 品种 | 假鳞茎长 | 茎粗 | 植株高 | 植株宽 | 叶长 | 叶宽 |
| 黄金富贵(59.67±3.06)a | (15.38±0.83) ab (392.33±1.53)a | (181.73±4.05) f | (122.67±24.85) bc (39.57±0.93)fg | ||||
| 2 | 东方小姐(41.38±7.42)c | (16.93±0.99) ab (267.67±82.80) b | (250.00±35.76) bc | (132.00±26.06) abc (39.03±5.25) fg | |||
| 3 | 贵妃 | (51.00±3.61) b | (18.80±3.40) ab | (276.67±7.77) b | (262.00±22.72) ab | (161.13±3.78)a | (46.70±2.52) def |
| 4 | 誓言 | (46.10±3.00) bc (19.10±0.61) a | (336.67±25.03) ab (297.67±11.24) a | (163.13±11.13) a | (66.10±1.35) a | ||
| 5 | 红玫瑰 | (28.67±4.16) def (18.33±2.90) ab | (238.67±29.30)b | (212.00±14.00) cdef (146.00±10.15) ab | (48.60±4.76) cde | ||
| 6 | 水晶 | (21.33±4.04)f( | (14.10±0.30) ab | (152.33±15.95)c | (110.00±6.08g | (86.33±8.08) d | (39.20±5.63)fg |
| 7 | 红金珠 | (33.33±1.53)d( | (16.23±0.15) ab | (254.67±3.21) b | (235.00±2.65) bcde | (137.33±1.53) abc( | (51.87±0.50) bcd |
| 8 | 朝日 | (25.00±2.00) ef (14.20±2.78) ab | (237.67±20.26) b | (224.00±10.54) bcdef (139.67±18.01) abc (35.47±4.10) gh | |||
| 9 | 泼墨石斛(23.00±1.73)ef(15.10±5.34)ab | (129.33±61.33)c | (190.67±28.87) ef | (104.67±8.74) cd | (28.27±7.21) i | ||
| 10 | 小男孩 | (24.00±1.00) ef (17.15±1.25) ab | (138.33±19.66)c | (197.33±24.95) def | (116.73±9.92) bc | (44.43±2.28) def | |
| 11 | 胭脂 | (57.40±1.25) a | (18.19±0.07) ab | (329.00±1.00) ab | (236.67±8.14) bcd | (150.67±1.53) ab | (57.00±1.00) b |
| 12 | 红霞 | (42.40±3.42) c | (15.68±4.27) ab | (277.67±20.11) b | (266.33±22.01) ab | (163.73±6.51) a | (44.00±1.23) def |
| 13 | 宝石 | (31.00±7.94) de (12.37±1.50) ab | (263.33±84.13)b | (233.33±8.96)bcde | (132.53±20.58) abc (30.77±0.93)hi | ||
| 14 | 彩霞 | (40.33±0.58)c | (17.17±1.85) ab | (238.00±7.81) b | (261.33±10.21) ab | (131.67±8.43) abc | 54.47±0.99) bc |
| 15 | 金如意 | (42.00±1.00) c | (11.87±0.15)c | (244.00±1.00) b | (223.00±2.00)bcdef | (117.00±2.00) bc | 43.47±0.49) ef |
2.2不同石斛兰品种植株花朵大小比较
石斛兰花朵性状是商品化选择的一个关键因素。表3可知,15个石斛兰品种花朵长度为 41.83~ 78.47~mm ,最长的品种为誓言,显著高于其他品种,最短者为泼墨石斛,其次为红金珠、小男孩;花朵宽度为 48.30~85.13~mm ,誓言最大,东方小姐第2,为 74.40~mm ,显著高于其他品种,泼墨石斛最小;花朵中萼片长度为 24.27~42.27\mm ,誓言最大,显著大于其他品种,其次为东方小姐、贵妃、红霞,红金珠最小;花朵中萼片宽度为
11.80~16.80\;~mm ,誓言最大,泼墨石斛最小,二者具有显著性差异,与其他品种差异不显著;花朵侧萼片长度为 24.67~42.17\mm ,誓言最大,显著大于其他品种,小男孩最小,其次为朝日、红金珠、泼墨石斛;花朵侧萼片宽度为 12.40~19.53~mm ,金如意最大,其次为东方小姐、誓言、黄金富贵,泼墨石斛最小;花朵唇瓣长为 20.77~38.93~mm ,誓言最大,其次为贵妃、东方小姐,显著高于其他品种,小男孩最小;唇瓣宽度为 12.03~36.77\mm ,誓言最大,泼墨石斛最小。

